pigeon Brazilian Journal of Genetics, 20, 4, 663-665 (1997)
(Revista Brasileira de Genética)
POINT OF VIEW
Dominance, codominance and epistasispigeons
By Wilmer J. Miller
My brief discussion on Dominance in the Brazilian Journal of Genetics 19, 4 (1996) p. 677 was designed to make people think about it and its various inconsistent usages. No one asked for a definition that I liked. One definition I use is couched in terms of the character: dominant - a character which we "see" or other wise detect, and which masks, hides, or completely obscures the expression of an alternative form of that character. (The word character may be replaced by the word difference for the same meaning).
This definition does not require the alternative form to be allelic. Thus, alternative forms such as black and white plumage color in chickens can be controlled by non-alleles (see the above reference). Defining a dominant gene does require the alternative form to be controlled by an allele, and should have much the same usage otherwise: a dominant gene controls a character (or difference) which masks, hides , or completely obscures the expression of an alternaltive form controlled by a recessive allele.
Thus, any intermediate form of dominance (which was not noted by Mendel) elicited a "hodgepodge" of definitions (often merely inferred) and names of the condition which are mostly contradictory between textbooks, confusing not only the students, but also the teachers, and perhaps the researchers! These names included: codominance, intermediate dominance, partial dominance, incomplete dominance, semidominance, transdominance, mosaic dominance, imperfect dominance, egalitarian dominance, lack of dominance, and absence of dominance (see Miller, W. J. and Hollander, W. F., 1995, Three neglected advance in classical genetics, BioScience 45: 98-104).
The definitions given or inferred for the intermediate form of dominance usually require knowing the exact function of the gene. This is very seldom known for most characters at present. All the "codominant" usages actually show or imply 3 phenotypes for 2 codominant alleles for the monohybrids genotypes in diploids. Why not use this consistency among names and their usages as the single criterion and use only one name? I prefer the shortest codominance.
So, for beginning students, as well as teachers and researchers, all genes fit one of 3 categories: dominant, codominant, or recessive. When more sophisticated nuances implying different functions are desired, then other names can be used with a definition (please) for this next level of usage. Other complexities such as reduced penetrance have been discussed in Miller and Hollander, Ibid.
The term Epistasis also has been used loosely or variously in textbooks of genetics and needs tightening. The usual definition one finds is: EPISTASIS- the masking effect of one gene pair over another gene pair. So epistasis/hypostasis is the same kind of relationship between gene pairs as dominance/recessiveness is within a gene pair. This seems simple and straightforward enough. But I object. A new term should define something characteristically different. It should separate one class of genes from another in a distinctive fashion. I contend that this definition is characteristic of nearly all genes* - not a unique condition. It is merely a further characterization of the action of most genes.
The last good discussion of epistasis I have noted is that by W. F. Hollander, 1955, Epistasis and hypostasis, J. Hered. 46: 222-225. Dr. Hollander traces the history of the term in its convolutions and mutations, for example from Bateson’s 1907 "piling on concept" of epistasis, side-tracked by Fisher’s 1918 zig-zag of epistacy (deviation from linearity), through Snyder’s 1940 "modernization" to a masking action rather than "latency". Evidently some prefer the use of the term interaction "qualified by other suitable adjectives". But the term epistasis remains in use.
I prefer to add one word, mutant, twice to the usual definition, i.e., EPISTASIS is the masking effect of one mutant gene pair over another mutant gene pair. This makes epistasis a special case of gene action, and therefore, a useful term. Hypostasis, of course, is the converse, and is applied to the gene pair hidden by the epistatic gene pair. (Most authors emphasize the term epistasis and seldom use the converse, hypostasis).
I have failed to note a description or listing in the textbooks or other sources of the 3 (or4) kinds of epistasis that exist. The term "blocking" fits their action.
THREE (or FOUR) TYPES OF EPISTASIS:
1. STRUCTURAL EPISTASIS - when a structure such as a hair can have polymorphism such as color. If the hair is missing (for example: hairless-possible mutant in mice and dogs), the color differences cannot be demonstrated., Another example is the black versus the white spines in cucumbers, Cucumis sativus, when a spineless mutant is present.
2. STERIC-HINDRANCE (structural in a different sense) - when a structure genetically present or absent gets in the way of an antibody reacting with its specific antigen, such as type A or B human bood groups "hiding" type Lewis-a from its specific antibody.
3. BLOCKING a sequential biochemical pathway - when the absence of one product prevents other products from being formed.
Using the usual textbook definition for epistasis...includes
4. CONVERSION - when the product of one gene is converted into another product by another gene, thereby masking the action/product of the previous genes. See Figures 1 and 2.
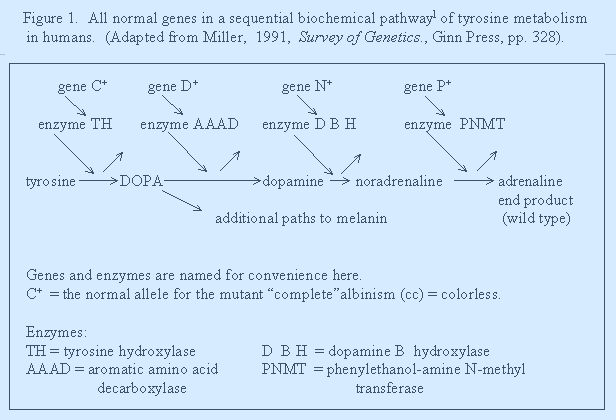
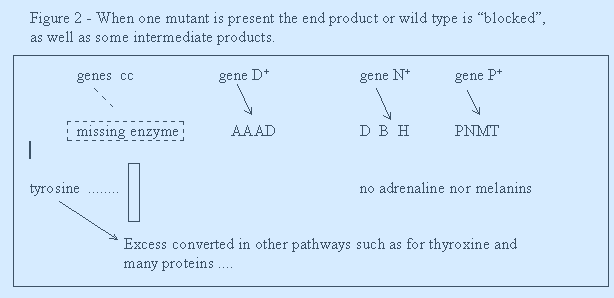
Is cc epistatic to ...P+...to wild type? If yes, then this reinforces the statement above * that nearly all genes are epistatic using the usual textbook definition. The single mutant action of cc is to block the normal gene action of D+ and of any genes contributing to the wild type of this character. That is, the cc mutant genotype is epistatic as well as that of the normal genes converting previous products. Thus, epistasis is expected of any mutant (!) as well as most normal genes as just shown above in figure 1.
In figure 3 the biosynthesis of aleurone is diagrammed with 2 side branches. Five mutants are known to interfere with the process. By the proposed new definition of epistasis we can list such action at the bottom of Figure 3. Additionally, mutant pr pr yields red aleurone by adding OH to the B ring of chalcone at the 3’carbon, The site of action of two other mutants c, white, and r, white, is unknown and their normal alleles may be regulatory.
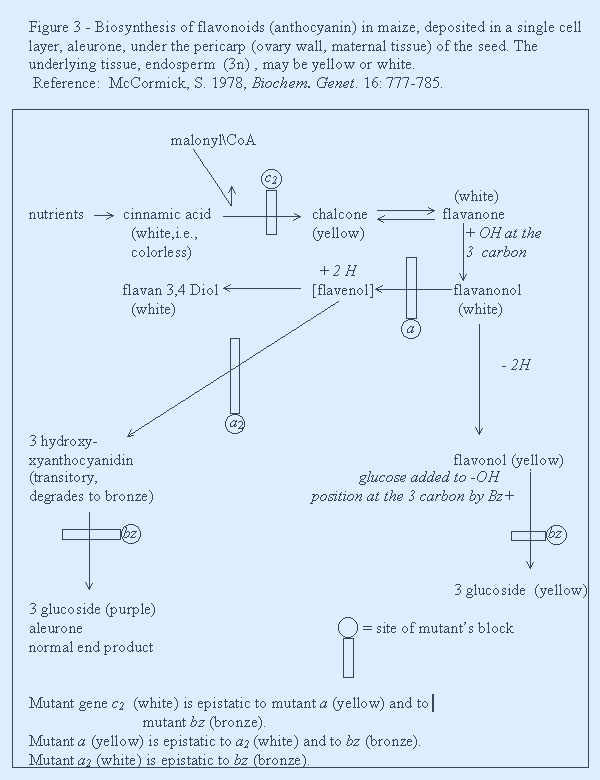
I think I have shown how the term epistasis is used rather loosely, Its use needs tightening! I have offered a new definition that I consider useful. Do you have a better one?
Shall we make epistasis a special case, or use it as an attribute of most genes? The readers can decide, but usage will dominate.