
9. X-Pigeons |
||||
~ PIGEON COLORS 2 ~
 Let's
recap what we've learned so far. According to Dr. Cryberg, pigeons
have the biochemical capacity to make at least three melanin pigments
in their feathers which are red, black, and brown. They also make
two non-melanin pigments that are found in their eyes which are pearl
and orange. The relationship between the melanin pigments and these
non-melanin pigments found in eyes are not understood yet. The eye
pigments are very different chemically but seem to be produced by
melonocytes. Pigeons also capable of making some lipichrome type
of pigment that is deposited in the legs making them red. Lipochromes
have nothing to do with melanin or melonocytes.
Let's
recap what we've learned so far. According to Dr. Cryberg, pigeons
have the biochemical capacity to make at least three melanin pigments
in their feathers which are red, black, and brown. They also make
two non-melanin pigments that are found in their eyes which are pearl
and orange. The relationship between the melanin pigments and these
non-melanin pigments found in eyes are not understood yet. The eye
pigments are very different chemically but seem to be produced by
melonocytes. Pigeons also capable of making some lipichrome type
of pigment that is deposited in the legs making them red. Lipochromes
have nothing to do with melanin or melonocytes.
The pigment study of feathers conducted by Axel Sell at al., after the Hollander Era, clearly proves a pigeon of any color (ash-red, recessive red, indigo, brown, etc.) is able to produce three pigments on all of their feathers; brown, black, and red; but this doesn’t mean they are the three so-called 3 base colors. Most literature about pigeon genetics start teaching the so-called b locus mutations first, where ash-red (red pigment) and brown (brown pigment) are alleles, and the wild-type gene at this locus (black pigment) in that allelic series codes for blue/black phenotypes.
Unfortunately, this method of teaching pigeon genetics leads readers to believe ash-red and brown are more important genes than the rest of the mutated genes found in other chromosomes. Hollander never talked about the so-called base colors, and it is still a mystery why some people in the past decided to claim there are 3 base colors in pigeons. The base, or wild-type, in pigeons or any other animal is a genetic concept. It is a phenotype we get when the most common genes found in the wild birds are all present. Therefore, there can only be one base color for pigeons; it is the black pigment with the appropriate modifiers to make a blue bar phenotype. Always remember that every single gene that makes the wild-type must be present and function properly for us to see all the characteristics of a wild-type phenotype. When one of them is not functioning correctly, and that gene happens to be affecting the phenotype of the bird, then we will no longer have a wild-type pigeon. Therefore, no genes are more special than others. There are genes on the sex chromosome that are part of the genes needed to make black pigment, but they are no more important than many other autosomal genes which also help making black pigments (recessive red, indigo, recessive white, dominant opal, etc.). We do not know how many actual locus points there are on a chromosome yet. But it is imperative to understand that all genes that somehow alter the color and pattern of the bird from our standard blue bar phenotype are as important as the ash-red and brown mutations.
 In
addition, anyone who thinks that the wild-type gene at b locus is the
only gene that makes
black pigment all by itself simply misunderstands how
genes work together. Many of us don’t seem to understand
that our wild-type, blue/black in that allelic series, is not a
phenotype
produced by a single gene only from that locus. In other words,
if the gene at the b locus is not brown or ash-red, it does NOT
mean we will automatically get a blue/black phenotype; because
every gene responsible for the wild-type phenotype in every other
locus and in every other chromosome “must” also be
coding for wild-type genes for us to be able to observe the wild-type
phenotype.
In
addition, anyone who thinks that the wild-type gene at b locus is the
only gene that makes
black pigment all by itself simply misunderstands how
genes work together. Many of us don’t seem to understand
that our wild-type, blue/black in that allelic series, is not a
phenotype
produced by a single gene only from that locus. In other words,
if the gene at the b locus is not brown or ash-red, it does NOT
mean we will automatically get a blue/black phenotype; because
every gene responsible for the wild-type phenotype in every other
locus and in every other chromosome “must” also be
coding for wild-type genes for us to be able to observe the wild-type
phenotype.
If you break any of the wild-type genes, you will not get a blue/black phenotype anymore. Indigo, recessive red, recessive white, albino, almond, dilute, reduced, grizzle, ice, recessive opal, dominant opal, and the list goes on. Thus, to think the wild-type gene at the b-locus solely responsible to produce blue/black phenotype simply not true at all, and will be confusing to people trying to understand pigeon genetics. It is imperative to understand that ash-red and brown mutations are not a bit more important than other mutations and should not be considered as the "main" or "base colors". Therefore, to categorize some genes as base colors is a very disingenuous way of thinking in modern genetics.
To better understand this concept, consider this scenario. Let’s say we have a pigeon that has wild-type genes in every other loci but at the b locus, it has two copies of the brown genes (b//b). In this case, the phenotype should be a brown bar. If we remove the brown mutant genes and replace them with the wild-type genes, we would get our standard blue bar phenotype back. Now, consider applying the same scenario by replacing the wild-type genes with the mutations for the following loci one by one for the recessive red locus, the almond locus, the indigo locus, the recessive opal locus, the recessive white locus, and the dilute locus. If a bird has all the wild-type genes in every other loci but have a copy of a qualmond gene at the almond locus for example, that bird would no longer have the wild-type (blue bar) phenotype.
Now ask yourself, if the wild-type gene at the b locus was solely responsible for making the bird a so-called “blue-based-bird”, what is the role of other wild-type genes located in other loci? Why does a bird have to have wild-type genes in every color loci in order to show a wild-type phenotype?
Just like the mutant genes located at the b locus (ash-red and brown), when the wild-type gene at the almond locus or at the indigo locus is mutated, we can no longer get our wild-type phenotype. Thus, what the fanciers call "blue base", which is known as the “wild-type” in genetics community is produced by all the wild-type genes in every loci acting together, rather than just the wild-type gene at the b locus acting all by itself. It is therefore, inaccurate to consider the ash-red as one of the so-called "base colors", when the mutant gene at the indigo, at the almond locus, and at many other loci also changes the wild-type phenotype by altering feather colors. We need to recognize that the wild-type gene at the almond and indigo loci must also do something special to produce the wild-type phenotype just like the wild-type gene at the b locus does. We do not know exactly what these mutants on the other loci do, but it is clear they all do something. It is clear because mutants of those locations all lead to something besides a wild-type. Sure, the wild-type at b locus is important, but not a bit more important than the wild-type at any other loci. In case you are wondering if the wild-type genes are more special than the mutants and the answer is a clear NO. The wild-type genes are simply the most common gene set in the population or the gene set that was thought to be most common when the standard was set. Nothing more, nothing less.
Biochemistry of genetics
 A
man, who was known as "Doc", Willard F. Hollander,
1913-2004
(left picture, taken on December 3, 2000) developed
methodologies including improved genetic
diagramming, symbols, and using a "wild-type" or standard
type reference that makes generalizing and analyzing classical genetics
more clear and
accurate than anyone before him. Dr. Willard Hollander was an internationally
renowned geneticist who had a long and distinguished
career. Throughout his academic career
in genetics he pioneered new fields of study especially about pigeons
and turned his childhood hobby into a career and experiences that
still remains unmatched. He was especially noted for his work in
deciphering the complex Mendelian genetics of pigeons, and for his
understanding of the complex genetic basis of the wide array of colors,
feather shapes and other characteristics that delight pigeon fanciers.
A
man, who was known as "Doc", Willard F. Hollander,
1913-2004
(left picture, taken on December 3, 2000) developed
methodologies including improved genetic
diagramming, symbols, and using a "wild-type" or standard
type reference that makes generalizing and analyzing classical genetics
more clear and
accurate than anyone before him. Dr. Willard Hollander was an internationally
renowned geneticist who had a long and distinguished
career. Throughout his academic career
in genetics he pioneered new fields of study especially about pigeons
and turned his childhood hobby into a career and experiences that
still remains unmatched. He was especially noted for his work in
deciphering the complex Mendelian genetics of pigeons, and for his
understanding of the complex genetic basis of the wide array of colors,
feather shapes and other characteristics that delight pigeon fanciers.
When the word "gene" was
invented, we didn’t have any ideas what a gene was on the DNA.
Mendel learned in the 1860’s that
some characters were inherited as units, he simply called these
factors units of inheritance. In the early 1960’s when Hollander
taught entry level genetics courses at the Iowa State University, he
refused and
prohibited
his
students to use the word "gene". His argument was using the
word gave the impression that his students knew what they were talking
about. He pointed out that at that point in time no one had a clue
what a gene was in terms of its total chemical makeup. They only knew
very little about what made genes work. So, Hollander used language
like a unit trait difference instead of gene. It was perfectly easy
to learn classic Mendelian genetics without ever using the word gene.
Today, we still don't fully understand everything there is to know
about genes and DNA. Although most people throw the word around like
it was known and all the details worked out, the process of understanding
the details is just starting to have the fog lifted.

Until recently, the vast majority of our DNA was dismissed as junk, but now we know it is important - or so you might have read recently. In fact, it still appears likely that 85 to 95 per cent of our DNA is indeed useless. While many bits of DNA that do not code for proteins are turning out to have some function or other, this was predicted by some all along, and the overall proportion of our DNA with a proven function remains tiny. Once, it all seemed so beautifully simple when we thought Our DNA consisted of a set of recipes, or genes, for making proteins. We thought if we had identified them all and worked out what they do, we would be a long way towards understanding what makes us what we are. If only the human genome project was not a big shock that we have only around 23,500 genes - barely more than a nematode worm, but in many other ways our genome turned out to be dizzyingly complex, confusing and intimidating.
Nonetheless, with the help of technology, the rate of progress is so fast nowadays that we will probably learn as much in the next ten years as we have learned in all of history up to today. Biochemistry has let us approach the genetic problems at the atomic level, and since it has turned out that the general plan of the macromolecules involved is rather simple, it has been remarkably successful.
Nowadays, scientist will look at the DNA sequencing data and biochemistry data far more convincing than Mendelian breeding data. Breeding data is a rather coarse screen and only gives general answers. That is why today genetics does very little at all with breeding data because we can’t get detailed understanding of genes using only breeding data. Today, most of the experiments in genetics science is done at labs and in test tubes. Nowadays, professional journals are not interested in publishing anything that is based on simply breeding data. In fact most of what we find in journals has only pedigree data with no actual breeding experiments, or at most if breeding experiments are needed they are little more than a footnote.
In the first page, I talked about DNA (deoxyribonucleic acid), which is a long-chain molecule that contains the genetic material -- encoded hereditary characteristics in living organisms. Now, let’s take the wild-type and pigment topics to the next level and learn how genetics governs biochemistry. Biochemistry, sometimes called biological chemistry, is the study of chemical processes and chemical transformations in living organisms. Much of biochemistry deals with the structures, functions and interactions of cellular components such as proteins, carbohydrates, lipids, nucleic acids and other components. When we look at the biochemistry of a chromosome we should notice that it is not just DNA, but a structure organized by a complex mixture of structural proteins, sequence-specific proteins, enzymes, and small RNA's. These components work together to facilitate reactions like transcription of ribosomal RNA and mRNA, DNA replication, and homologous recombination. The RNA, (ribonucleic acid) is also a long-chain molecule just like the DNA. The function of RNA is to translate the genetic material stored in DNA, into protein structures. RNA essentially carries out the instructions of DNA. In most cases the words in the RNA are converted into a protein where each three letter word corresponds to one single amino acid in the protein chain. RNA plays a central role in the pathway from DNA to proteins. An organism's genetic information is encoded as a linear sequence of bases in the cell's DNA. During the process known as transcription, a RNA copy of a segment of DNA, or messenger RNA (mRNA), is made. This strand of RNA can then be read by a ribosome to form a protein. Therefore, the most important function of genes is to direct the production of protein molecules where the nucleic acid is the main genetic material.
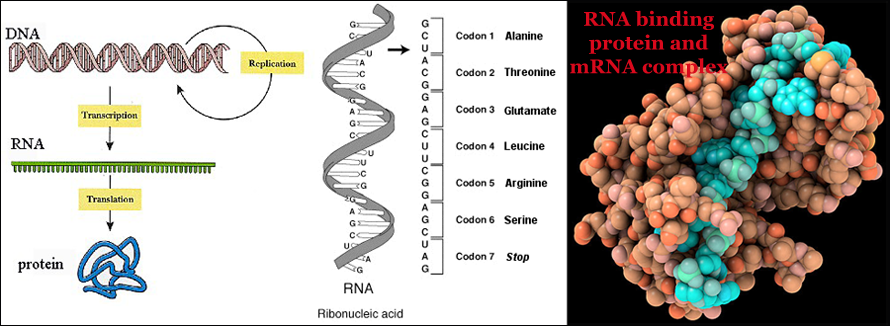
Let’s evaluate how do these three well known pigments (red, black, and brown) that pigeons are able to produce in their feathers controlled by dominant, co-dominant, and recessive genes. According to Dr. Cryberg, there is very limited number of things that genes are known to do chemically, and no gene directly makes pigments. Instead, a gene makes an RNA molecule and RNA molecule is usually translated into a protein. Biological pigments are substances produced by living organisms that have a color resulting from selective color absorption. Many biological structures, such as skin, eyes, fur, feather and hair contain pigments such as melanin in specialized cells called chromatophores. Melanin is a class of compounds that serves as a pigment with different structures responsible for black, brown, and yellowish/ reddish pigments in the feathers of pigeons. There are two classes of pigments: black and brown insoluble eumelanins, which are derived from aerobic oxidation of tyrosine in the presence of tyrosinase, and the alkali-soluble phaeomelanins which range from a yellow to red brown color, arising from the deviation of the eumelanin pathway through the intervention of cysteine and/or glutathione. Eumelanins are usually found in the skin and eyes. The primary function of pigments in plants is photosynthesis, which uses the green pigment chlorophyll along with several red and yellow pigments that help to capture as much light energy as possible. Other functions of pigments in plants include attracting insects to flowers to encourage Pollination. Pigmentation is used by many animals for protection, by means of camouflage, mimicry, or warning coloration. Some animals including fish, amphibians and cephalopods use pigmented chromatophores to provide camouflage that varies to match the background. Pigmentation is used in signaling between animals, such as in courtship and reproductive behavior. For example, some cephalopods use their chromatophores to communicate. Skin pigments such as melanin may protect tissues from sunburn by ultraviolet radiation.
When pigeons produce pigments in their feathers, proteins cause chemical reactions which ultimately cause some step in pigment formation to be speeded up or slowed down. Even though vast majority of the genes code for protein, we know of a very few genes that do not even make RNA, and only a few stop at RNA. More importantly, no wild-type genes or any mutations will ever be discovered to directly make pigments, because pigments are not proteins. All a gene does is dictate the order of the amino acids that are put into a given protein. If a gene mutates, the mutation puts an incorrect amino acid someplace in the protein. When this happens, some errors make little difference and the protein still works OK. But, we know that most errors result in problems of some kind. In the case of enzymes or proteins that bind to DNA and govern transcription errors that result in problems generally cause either 100% loss of function or near 100% loss of function.
Interactions
between recessive, dominant & co-dominant
mutations 
Mutations happen all the time and most of these mutations are recessive mutations. However, most of these mutations are eliminated from the gene pool in the wild because birds in wild do not inbreed, especially for number of generations. Therefore, newly mutated recessive genes do not get to be propagated, and hardly reach the homozygous state. Even in domesticated pigeon lofts, many pigeon breeders do not inbreed a lot, so that the new recessive mutant does not become homozygous and eventually gets eliminated. However, given enough time all of the recessive genes we know of and the new ones would show up and get incorporated in domestic lofts, if people did not cull them as fast as the mutants showed up. Pigeon breeders probably cull some new mutants from the hobby every year because new mutants do not fit the owner’s idea of what is proper and acceptable for his/her breed. On the other hand, most of the time new mutations are reported by average pigeon fanciers if and when they recognize something unusual. For instance lemon (extreme dilute) was first recognized and characterized in Barkel’s Homing Pigeons in South Africa. We have data to prove that totally unrelated Homers in Portugal also had the same mutation. We can even trace back in this case to the bird the mutation happened in an ash-red intense hen. Apparently she had a mutation at some point that ended up making the tissues in her ovary as she passed the mutant on to two different sons. Two independent tests show this mutant found in Portugal is phenotypically the same as Barkel's from South Africa. In United States, we have two additional less documented examples of what phenotypically looks the same as lemon mutations. It is perfectly possible that all four of these lemon mutations are actually different mutants if they do prove that all four be alleles. There are lots of ways to totally break any gene so there are lots of alleles that will give the same phenotype.
The lemon example given here, where we know of several cases what seems to be the same mutation has happened several times, is a good enough example to prove that mutations happen all the time. Furthermore, some mutations are found even in different families of pigeon species. We know that barless pattern mutation that we see in our pigeons (Columba livia) is also found in Wood Pigeon (Columba palumbus) which suggests that there is analogous mutation at the same spot on the chromosome in both Columba livia and Columba palumbus. Since pigeons are related evolutionarily, we'd expect major parts of their genomes to be identical. The same mutation in one spot in one species, would likely lead to an analogous (identical) mutation in another. We have extreme dilute in the ringneck dove and now we have extreme dilute (lemon) in pigeons and no one believes that extreme dilute was added to pigeons from the doves - rather there was the same mutation at the same spot in two different species. Modern science predicts that there was a common ancestor between ringneck doves (Streptopelia risoria) and rock pigeons (Columba livia) and this ancestry goes back to 25 to 30 million years. This number is easier to say than comprehend how long it took for both species to evolve from their common ancestor. For years we knew crossing ringneck doves with pigeons produced viable but not fertile hybrid offspring. We also knew both of these domesticated species had peak crests. Crossing crested dove with a crested pigeon produced crested hybrids, suggesting a similar genetic basis for this trait in both species. Thus, the common understanding is that both cr mutants found in these species break the functionality of the gene somehow and it does not matter how the gene functionality is broken since the result produces the same morphological effect on the phenotype in both species – head crest.
When a wild-type gene mutates to a recessive gene, it is an indication that the functionality of this wild-type gene is broken and the protein is no longer capable of performing its normal function. If only one of the copies of the gene is a recessive gene and the other one is still the wild-type (heterozygous state), then we normally cannot see phenotypic affects of the recessive genes. However, when both copies of the wild-type genes are mutated to recessive genes, then we can no longer have the wild-type phenotype; instead, we will see the effects of the recessive genes. When two of the recessive or dominant allele genes are present in the genotype (one copy of ember and one copy of recessive red, or one copy of lemon and one copy of dilute) a phenotype will seldom get intermediate phenotypes from alleles. However, we will get intermediate phenotypes all the time from non alleles. It is therefore wrong to draw a conclusion from intermediate phenotypes to compare if the genes are allelic or not. So, what should we expect to see when two of the recessive alleles are present in the genotype if they won’t give us intermediates? For instance, what should the phenotype be, if one copy of recessive red and one copy of ember are present at recessive red locus? The answer is ember because from the breeding results we know that ember is dominant to its allele recessive red. What will the phenotype be if one copy of dilute and one copy of lemon are present at the dilute locus for the cock birds? Dilute, because dilute is dominant to its allele lemon based on breeding results. If there is one copy of pale and one copy of dilute are present at the dilute locus for the cock birds we would get a pale phenotype. In all such cases the phenotype is simply that of the more dominant of the pair of alleles. Furthermore, if we have the technology to cut the recessive red locus out of the pigeon genome, we would never ever get a blue phenotype anymore. That bird would be recessive red. If we cut out the recessive white locus out of the genome, all our birds would be recessive white. This means that having two copies of the most broken recessive genes at one locus (recessive red, recessive white, lemon, etc.) is same as taking (cutting) this locus out of the genome because in both cases the final result will be the same. Understanding recessive genes are probably easier compared to understanding dominant and co-dominant genes but, there is something else I would like to point out about the recessive genes.
 We
say that recessive genes can only be expressed if and when they
are in homozygous state.
Well, it is not entirely true. Actually, it
is only
true if the recessive mutant is tested against the wild-type when no
other mutants are present. We rate the new mutant against the wild-type
phenotype as dominant, co-dominant or recessive so that everyone understands
the rating system and use it the same way to eliminate confusion. There
is absolutely nothing at all magical or particularly special about the
wild-type phenotype. It is simply the standard science picked. When
a mutant gene is said to be recessive, we should all automatically
know that it is recessive to the wild-type and wild-type only. Recall
that we can only test a mutant against the wild-type or to its alleles.
We
can
test
if ember is dominant or recessive to recessive red mutant, because
they are alleles. However, we cannot rate if recessive red is dominant
or recessive to recessive white mutant, because they are not alleles.
We don’t expect a recessive
gene to express in heterozygous state when we test it against
the wild-type. But, that really does not tell us how any recessive mutant
will act in the presence of other mutant(s). In fact, a great many
mutant genes, which we
say are recessive, will show up plainly in the phenotype when combined
with other mutants.
We
say that recessive genes can only be expressed if and when they
are in homozygous state.
Well, it is not entirely true. Actually, it
is only
true if the recessive mutant is tested against the wild-type when no
other mutants are present. We rate the new mutant against the wild-type
phenotype as dominant, co-dominant or recessive so that everyone understands
the rating system and use it the same way to eliminate confusion. There
is absolutely nothing at all magical or particularly special about the
wild-type phenotype. It is simply the standard science picked. When
a mutant gene is said to be recessive, we should all automatically
know that it is recessive to the wild-type and wild-type only. Recall
that we can only test a mutant against the wild-type or to its alleles.
We
can
test
if ember is dominant or recessive to recessive red mutant, because
they are alleles. However, we cannot rate if recessive red is dominant
or recessive to recessive white mutant, because they are not alleles.
We don’t expect a recessive
gene to express in heterozygous state when we test it against
the wild-type. But, that really does not tell us how any recessive mutant
will act in the presence of other mutant(s). In fact, a great many
mutant genes, which we
say are recessive, will show up plainly in the phenotype when combined
with other mutants.
Some bar pattern birds that are heterozygous barless have very narrow bars for instance, but some do not. A heterozygous recessive red will punch a poorly defined bronze bar right thru the black on such a bird. In addition, a hetero recessive red can become obvious on some ash reds and also in some indigo phenotypes. But, there are things besides recessive red that will give the same phenotypes on ash and indigo so just because you see some phenotype is not proof it is recessive red. A classic almond (picture above), which is thought to be one of the most attractive and desired phenotypes by pigeon breeders, is a phenotype caused by many mutant genes acting together. The formula for a classic almond cock is heterozygous stipper, heterozygous recessive red, homozygous T-pattern check, and homozygous kite bronze. If we take the recessive red out of this formula, the ground color on the bird simply is not bronzy enough to be a classic almond anymore. Therefore heterozygous recessive red is an essential ingredient on a classic almond phenotype. What we need to understand is, even though recessive genes can sometimes show in heterozygous state on some phenotypes depending on the presence or absence of other mutants in the genotype, that does not change the fact it is a recessive gene when rated against the recognized standard -- wild-type.
 Let’s
talk about how the recessive allelic series like gazzi-pencil-recessive
white, or recessive red-ember, or reduced-rubella, or dilute-pale-extreme
dilute (lemon) are rated against each other. At recessive red locus
for instance, if we have one of the genes broken, but have a working
copy of the wild-type gene, then the single wild-type gene will still
allow normal (wild-type phenotype) pigment production. The problem
occurs when both of the wild-type genes at this locus are broken.
We know that there are two broken versions of the wild-type gene
at this locus: recessive red and ember (picture
on right). The
most broken gene must be recessive red as it is the most recessive
of the series. Recessive red probably totally destroys the function
of the wild-type protein produced by the wild-type gene at recessive
red locus. However another recessive allele at this locus, ember,
probably destroys the function of the wild-type gene by 90% or 95%
where there is a tiny amount of function of the gene left. So, recessive
red will be the most recessive, ember will be dominant to recessive
red and wild-type at recessive red will be dominant to ember. At
the dilute locus wild-type is the most dominant, then comes pale,
dilute and lemon. So the order of dominance in this recessive allelic
series is: wild-type>pale>dilute>lemon. At the recessive
white locus the order is: wild-type>gazzi>pencil>recessive
white. At the reduced locus the order is: wild-type>rubella>reduced.
Let’s
talk about how the recessive allelic series like gazzi-pencil-recessive
white, or recessive red-ember, or reduced-rubella, or dilute-pale-extreme
dilute (lemon) are rated against each other. At recessive red locus
for instance, if we have one of the genes broken, but have a working
copy of the wild-type gene, then the single wild-type gene will still
allow normal (wild-type phenotype) pigment production. The problem
occurs when both of the wild-type genes at this locus are broken.
We know that there are two broken versions of the wild-type gene
at this locus: recessive red and ember (picture
on right). The
most broken gene must be recessive red as it is the most recessive
of the series. Recessive red probably totally destroys the function
of the wild-type protein produced by the wild-type gene at recessive
red locus. However another recessive allele at this locus, ember,
probably destroys the function of the wild-type gene by 90% or 95%
where there is a tiny amount of function of the gene left. So, recessive
red will be the most recessive, ember will be dominant to recessive
red and wild-type at recessive red will be dominant to ember. At
the dilute locus wild-type is the most dominant, then comes pale,
dilute and lemon. So the order of dominance in this recessive allelic
series is: wild-type>pale>dilute>lemon. At the recessive
white locus the order is: wild-type>gazzi>pencil>recessive
white. At the reduced locus the order is: wild-type>rubella>reduced.
So, recessive white, reduced, lemon, and recessive red are the most broken genes at their recessive allelic series. We can conclude that recessive genes simply result from taking some gene that does something important in making what we call blue phenotype and busting that gene so it no longer functions or at best functions poorly.
The sex-linked brown mutation is the broken or non-functional version of the wild-type gene at what is known as the b locus. If the wild-type gene at the b locus is inactive, non-functional, or broken, we can no longer get our wild-type (blue/black) phenotype – instead we would get a brown phenotype. When the wild-type genes at the brown locus are not functioning correctly anymore, the bird will still be able to make lots of pigment in its feathers. However, the pigments would just be lots of brown and limited amounts of red. So, if the wild-type genes at the brown locus did not exist at all, blue/black would not be known as the wild-type gene at the brown locus as what turns brown pigment into black pigment. This is the exact reason why we can’t call the wild-type gene at brown locus the “blue gene”. Because if we did that, we would have to call the wild-type gene at the recessive red locus also a blue gene, since it directly affects the phenotype of the bird and allows us to get something other than a blue/black bird. According to Dr. Cryberg Nearly all recessives allelic series are going to work like this where a mutation have destroyed the function of some important protein. Same thing is happening with lemon, dilute and pale where lemon 100% destroys the function of the wild-type gene at the dilute locus. Dilute is likely caused by wild-type gene’s loss of function by 95% and pale is caused by 90% functional loss. Understand that this series would look just the same if the % destroyed numbers were 100% for lemon, 70% for dilute and 30% for pale. However, it is hard to destroy the function by 70% and 30% and therefore using 95% and 90% works better as an explanation.
 Dominant
genes on the other hand, have lots of other options where there are
simply a ton
of things that can be happening and we cannot
even guess
which is correct. Recessives are much easier to understand
as they simply are the net result of massive loss of gene function. With dominants
we do
not even know if there is a functional wild-type gene doing
anything at that locus. Wild-type could even be a deleted gene and there is nothing
there coding for protein. Also a dominant mutant could simply
retain
a functional gene that came from some far ancestor. We
know that every wild-type gene must be present and function correctly for
us to
be able
to see the wild-type phenotype. When a wild-type gene mutates
to a dominant or a co-dominant gene, then that gene’s products such as
RNA and protein are still considered as functional – the gene products
act on the organism but change it in some way. For some dominant genes, it
is entirely possible that the wild-type version produces a protein that does
not
work at
all while the mutant works just fine. Suppose for instance
that most wild
pigeons were reds. Then the wild-type would be defined
as red. But an occasional wild bird (or domestic) shows up as a blue phenotype
where
testing would show that the blue form is dominant to wild-type
(red in this analogy), because we know that there is chemistry that is required
to avoid making red pigment when red is not in homozygous state.
Dominant
genes on the other hand, have lots of other options where there are
simply a ton
of things that can be happening and we cannot
even guess
which is correct. Recessives are much easier to understand
as they simply are the net result of massive loss of gene function. With dominants
we do
not even know if there is a functional wild-type gene doing
anything at that locus. Wild-type could even be a deleted gene and there is nothing
there coding for protein. Also a dominant mutant could simply
retain
a functional gene that came from some far ancestor. We
know that every wild-type gene must be present and function correctly for
us to
be able
to see the wild-type phenotype. When a wild-type gene mutates
to a dominant or a co-dominant gene, then that gene’s products such as
RNA and protein are still considered as functional – the gene products
act on the organism but change it in some way. For some dominant genes, it
is entirely possible that the wild-type version produces a protein that does
not
work at
all while the mutant works just fine. Suppose for instance
that most wild
pigeons were reds. Then the wild-type would be defined
as red. But an occasional wild bird (or domestic) shows up as a blue phenotype
where
testing would show that the blue form is dominant to wild-type
(red in this analogy), because we know that there is chemistry that is required
to avoid making red pigment when red is not in homozygous state.
We could say that the wild-type gene at the spread locus as we know it may not be doing anything at all to contribute pigment production; however the mutated version (spread) does something to change the phenotype. In addition, there is a possibility that the spread mutant produces a protein that binds to DNA and blocks some function with the result that we make a black bird instead of a blue. But, wild-type could make a protein that serves some totally different function. Maybe the wild-type protein is involved in making collagen. The mutated (spread) form of the protein might still work ok in its job to make collagen but it just happens to also bind to DNA now and regulate DNA function. So, it is doing two unrelated jobs and as it only takes a single mutant to bind to the DNA that mutant will be dominant.
Ash-red, Indigo, and Spread are examples of some of the dominant mutations in pigeon genetics. That is a hint that the wild-type form does much less than the mutant. The dominant mutant could simply be making some product that is antagonistic to some other genes functioning and thus blocking that normal gene from doing its normal job. In such a case, the wild-type is just sitting there doing nothing while the mutant is the form doing something. It is possible that in some ancestor of pigeons it was doing something which may have had more survival value then doing nothing and today the situation may have been reversed. Consider recessive opal which comes in different forms: The most recessive form has been properly named cherry by Steve Souza, who provided data to show that it is recessive to the blue form. This most recessive allele of the recessive opal is the most broken of course, and it looks a great deal like ash-red or homozygous indigo. According to Dr. Cryberg, a cherry bird in bar pattern has red bars, a very light colored tail, and a negative tail bar, just like the ash-red. So, we can tell that the wild-type gene at recessive opal does something real important in making a normal blue phenotype. We know that ash-red is a dominant and indigo is a co-dominant gene. Ash-red looks a great deal like cherry as does the homozygous indigo. So, both the ash red and indigo mutants produce some protein that somehow stops wild-type at recessive opal from doing its normal function. They both might bind to DNA and prevent wild-type at recessive opal from being transcribed into protein. They could bind with the protein wild-type at recessive opal produces and thus render the protein ineffective. They could refold the protein again making it ineffective. They could bind with some starting material that wild type at recessive opal protein needs to do its job. Therefore it is very hard to understand and explain dominant and co-dominant genes as we don’t know exactly what might be going on with the dominant mutant.
There is another condition that differs from recessive, dominant, and co-dominant genes. Some mutations like recessive red, recessive opal, and recessive white, and albino, alters the pigment production. So, when these mutations present themselves in the phenotype in homozygous state, they conceal the other mutations like ash-red, indigo, brown, blue/black, etc. This condition is known as epistasis, where one gene suppresses the effects of another non-allelic gene. In other words, when two genes are present in the genotype but only one of them is fully expressed in the phenotype, then the gene that is expressed is said to be epistatic. So far, there isn’t any scientific research done to see what chemical changes takes place to change for example a red pigment found in ash-red mutation to appear as recessive red on the phonotype. We do not know if they change the feather structure completely and turn on/off the pigment production all together. What we do know is that the pigment found in recessive red and ash-red is the same pigment, known as the phenomelanin in chemistry, but the way they are laid down in the feathers changes the appearance, giving us two distinct phenotypes. For instance, if we have a homozygous ash-red cock (BA//BA) or a hemizygous ash-red hen (BA//.) and if these birds happen to have two copies of recessive red mutations (e//e) then these birds will still produce red pigments in their feathers. However, instead of showing ash-red phenotype, they will instead be recessive red, and the recessive red mutation will also conceal the pattern of the bird.
 Let me give you a scenario
to explain all the epistatic mutations we have in pigeons up to this
point. Suppose we have a bird that is homozygous albino (al//al),
homozygous recessive red (e//e),
homozygous
spread (S//S),
homozygous recessive white (zwh//zwh),
and is wild-type in every other loci. Notice that all the mutations
except “spread” are
recessive to wild-type and therefore they can only be expressed in
homozygous state.
Let me give you a scenario
to explain all the epistatic mutations we have in pigeons up to this
point. Suppose we have a bird that is homozygous albino (al//al),
homozygous recessive red (e//e),
homozygous
spread (S//S),
homozygous recessive white (zwh//zwh),
and is wild-type in every other loci. Notice that all the mutations
except “spread” are
recessive to wild-type and therefore they can only be expressed in
homozygous state.
In this example, with all these mutations present in its genetoype and function correctly, this particular bird will be an albino with pink eyes on its phenotype. Why? Because albino is epistatic to “all” known colors and patterns. If we take the albino genes out of the equation and replace the albino locus with the wild-type genes, then this bird would now be a recessive white in its phenotype without the pink eyes (more than likely with bull eyes). If we take the recessive white genes out of the equation and replace the recessive white locus with the wild-type genes, this bird would then have a recessive red phenotype. If we take the recessive red genes out of the recessive red locus, and replace them with the wild-type genes, then this bird now would have a spread black phenotype. Lastly, if we take out the spread mutation and replace that locus with wild-type genes, then this bird would now have wild-type genes in every loci and would present itself as our standard blue bar, wild-type.
Therefore, in pigeons, we can say that spread is epistatic to all known patterns (T-check, Check, Bar and Barless). Recessive red is epistatic to all patterns and most color mutants (except recessive white, and albino). Recessive white is epistatic to all know colors and patterns (except albino) and finally albino is epistatic to “all” known color and pattern mutations.
Recessive red - e
The mutant gene we call recessive red does not make red pigment at all. This mutant simply stops the formation of black pigment. With the black pathway blocked the biochemical machinery is forced to take a sideways reaction that makes red pigment after numerous other steps governed by numerous other genes. The recessive red only present itself in homozygous state (e//e) when tested against wild-type. Otherwise, in heterozygous state (e//+), a pigeon just carries that gene in its genotype but does not show it in its phenotype. In heterozygous state, the wild-type pigment at the recessive red locus is able to produce the black pigment even with a single copy. When the locus for recessive red has two copies of the mutated non-functional genes (e//e), the bird is not able to produce the black pigments without the normal functioning wild-type genes and this bird would present itself as a recessive red; unless the same bird is homozygous for recessive white or homozygous for albino. Therefore, we could claim that the function of the wild-type gene at the recessive red locus must not be just producing the normal amount of black pigment, but also help distributing that pigment evenly to all the feathers where we normally expect to see black pigments, including bars and sub-terminal tail bands. Since two copies of recessive red mutation at this locus stops the production and distribution of the black pigment, we get ourselves a nice recessive red bird covering all other pigments and pattern of the bird. If you ever produce a recessive red offspring, it proves that both parents are heterozygous for recessive red mutation (e//+).
 Recessive
red is an epistatic mutant to rest of the color mutation
(except recessive white, and albino) and patterns. Therefore, if you
have a recessive red bird, and have no clue about its pedigree, it
can be carrying many mutant genes that were masked by recessive red
mutation. However, it seems even a single copy of spread gene carried
in the genotype increases the quality and the richness of recessive
red phenotype. Any fleck results in a feather from extra pigment deposited
versus the surrounding areas by definition. Flecking happens in lots
of different gene combos; some a lot more than others. Some
recessive reds seem to have near black areas on the feathers. Thus,
it seems there is not anything unique at all about flecking in an ash
red other than the color contrast makes them easy to see.
Recessive
red is an epistatic mutant to rest of the color mutation
(except recessive white, and albino) and patterns. Therefore, if you
have a recessive red bird, and have no clue about its pedigree, it
can be carrying many mutant genes that were masked by recessive red
mutation. However, it seems even a single copy of spread gene carried
in the genotype increases the quality and the richness of recessive
red phenotype. Any fleck results in a feather from extra pigment deposited
versus the surrounding areas by definition. Flecking happens in lots
of different gene combos; some a lot more than others. Some
recessive reds seem to have near black areas on the feathers. Thus,
it seems there is not anything unique at all about flecking in an ash
red other than the color contrast makes them easy to see.
Under a microscope, the "red" pigment in ash-red and recessive red is still "red", but how it's laid down is totally different with both mutations. The fact of the matter is we don’t know exactly where in the pigment forming process the mutation interacts. When we have two different mutations interacting with the same pigment base (ash-red, and recessive red), and yet producing different results and being the result of mutations on totally different chromosomes is a mystery. What we do know is that recessive red is neither sex-linked nor dominant. Instead, it is a recessive autosomal mutation. Recessive red cannot be mistaken for spread ash-red because in recessive red the flights and the tail feathers are a solid red and do not show the typical light ash colors. This is also true for unimproved recessive reds that shows lighter blue/black color in the tail feathers.
Wild-type can be epistatic to some mutant; however, this mutant can show only if some other mutant is present. An example would be white sides on recessive red. A recessive red white sides bird is a self red bird with white wing shields. Yet, if you simply replace the recessive red mutant with wild-type at that locus you will get a self blue bird with no white showing even when the mutant for white sides is homozygous. So wild-type is said to be epistatic to the white side mutant. White side is neither dominant, nor recessive, nor codominant. It simply does not show on an otherwise wild-type pigeon. There are other known examples where wild-type is epistatic to some mutant. The recessive red and recessive white pictures in this page are submitted by Vahe' D'Ala. www.falconlofts.com
Recessive white - zwh
When recessive white is
in homozygous state (zwh//zwh),
what happens is that either the melanin is never produced
or it doesn't get placed into the feather
of the
bird. Melanin is the pigment which is normally in a pigeon's feathers
providing the colors we see depending on its shape and amount. Feathers
of the bird grow normally, but, without melanin available to color
it — only a white feather is produced. Since
in homozygous state recessive white does not allow or make minimum
amounts of pigments which are not visible to naked eye, the function
of the wild-type gene at the recessive white locus must be to make
tyrosinase or provide tyrosine to the pigment synthesis point. We
actually see the feather as a white color because light hits the
feather and bounces back
to our eyes. Since none of the various light rays are absorbed by
the melanin, all of them are bounced back and we see the combination
of them as white. Recessive white plumage is often confused with albino
mutation although they are two separate mutations caused by different
genes. Separating these two phenotypes is actually quite easy when
we take a close look at their eyes where albino pigeons have pink
eyes and recessive white birds have bull eyes. 
The bull eye is generally seen in recessive white and piebald phenotypes. All recessive white pigeons have bull eyes, white feathers, white beak, white skin and white nails. It's because just like the eye, rest of the phenotype genes for feather, beak, skin, and nail the melanin production is turned off at the surface level. Because there is no melanin on the surface level, we see white. The bull eye therefore hides two additional genetic eye pigments under it, and does not allow us to see it since the melanin production on the outer iris of the eye is missing or not functioning. Therefore, we can claim that bull eye is epistatic to orange and white eyes.
Remember, recessive traits need two genes carrying that information to present itself fully on the phenotype of the bird. If a bird is homozygous for recessive white (zwh//zwh) then, no matter what color it would normally be, in every case, such a bird will be a white pigeon. That means a white bird can literally be almost anything genetically underneath those white feathers. It could be a blue-bar, a red check, a recessive red, a reduced T-pattern blue check, spread indigo, etc. The only way to find out what color a recessive white actually carries underneath those white feathers is to mate it with a blue bar, assuming that blue bar is not heterozygous for recessive white. All the offspring from this mating will be able to produce melanin in the offspring’s feathers which we can compare the mutation producing the pigments to our wild-type standard.
Recessive opal - o
Just like recessive red and recessive white, the recessive opal is also an autosomal recessive autosomal mutation. This mutation was recognized as something unique and its inheritance worked out by W.F. Hollander in the 1930's. Recessive opal is very common in racing pigeons and there's a good amount of variation in coloring from bird to bird. Most have lightened tail and flight feathers. Their bars and checks are pinkish-bronze-grayish. A few birds are near-perfect mimics for Ash-reds. Ash-red gives a phenotype quite like homozygous red phase recessive opal. These are the ones which confuse people who believe they've produced ash-reds from a pair of blue birds. In reality, they have produced recessive opal that mimics the ash red. Because of this, sometimes they are even confused for reduced and dominant opal birds but they are two very separate mutations and each inherits independently of the other.
Dilute - d
We get cream bar (ash-yellow) from ash-red, dun from black, silver-dun from blue, recessive yellow from recessive red, and khaki from brown by a mutation called dilute. The dilute gene is located in a different locus (different forms of a gene at a particular physical location on a chromosome) and therefore inherited independently. Dilute is a sex-linked recessive mutation. Dilute is self-descriptive and it does what it says -- it dilutes the original pigment color giving us a faded version of that same color. Dilute gene is a recessive gene which can be carried but not shown for the cock birds, but if the hen has a single dilute gene in her only functional sex-chromosome, donated by her father, she will be showing the dilute factor in her phenotype. In order to raise a dilute cock, both parents must donate the gene to their offspring and make the bird homozygous for dilute (d//d). Hemizygous hens (d//.) can be raised showing dilute if her father carries a single factor (heterozygous for dilute) and if he donates it to his daughter. This also means that if a cock bird is homozygous for dilute, then all the female offspring from this cock will be diluted. In other words, if you ever produce a diluted bird form non-dilute parents, it means this offspring is a hen and her father classified as a heterozygous for dilute.
The dilute phenotype can get a little tricky because there is one rare exception to the rule that if any time two normal intensity birds produce a dilute that dilute will be a hen and short-downed. There is a rare gene called “pink eyed dilute” that is autosomal and recessive. So, in theory two normal looking birds that are heterozygous for pink eye dilute gene can produce either sex young as dilutes. Even though it is a very rare condition, when it happens this mutation in homozygous state causes poor eyesight and the adults have pink eyes.
The effects of dilute can sometimes create confusion describing the phenotype caused by other mutations. For instance, dilute version of blue/black known as the silver dun can often be confused with brown mutation. One way to separate them is look at them couple of days after hatching. Dilute birds are short-downed appearing naked compare to brown birds. However, short-downed can be caused by other mutations; it is therefore important to note that although all diluted birds are short-downed, it doesn’t mean every short-downed squab is a dilute. Also, brown pigmented birds known to have what we call “false pearl eyes”. Again, this doesn’t mean every pearl eyed bird will be brown, but if you have a brown looking bird, and if it doesn’t have pearl eyes, you can be sure that it is not a brown bird.
Remember that for a cock bird to be a dilute bird, he must be homozygous for dilute. I am not sure if it was a coincidence or most diluted birds are hens, but I noticed after I posted the pictures that the first row of the following pictures are all cocks, and the second row showing the dilution is all hens. Click on each picture to make it bigger. In the pictures below, the dun female is actually the daughter of the black bird above her. How? Because this black bird is heterozygous for dilute. This means that chances of this cock bird producing a diluted hen offspring is 50%.
) |
) |
) |
) |
) |
) |
) |
) |
According to a study done by Haase E., Ito S., Sell A., and Wakamatsu K, (1992) “Melanin Concentrations in Feathers from Wild and Domestic Pigeons,” Journal of Heredity, the term “dilution factor” suggests that the effect of this factor on the color of pigeons is accomplished by a decrease of pigment concentrations. In an early attempt to quantify the effects of this locus, Lloyd-Jones (1915) compared red with yellow and black with dun birds. Lloyd-Jones concluded that red and black pigments are probably affected in a constant manner and that their concentrations are increased three times in intense vs. dilute colored birds. The results of “Concentrations in Feathers from Wild and Domestic Pigeons” study shows that in the eumelanic bird, dilute mutation decreases the eumelanin concentration whereas it does not affect pheomelanin levels. In this study authors compared 9 black vs. 12 dun, and 13 red vs. 14 yellow birds. In the pheomelanic homozygous recessive red (e//e) birds, again eumelanin is decreased three times, but in this genetic context pheomelanin is reduced about five times. Therefore, the effects of a dilute mutation cannot simply be described by a single dilution index. As in the case of spread factor (S), the effects of dilute (d) on eumelanin and pheomelanin concentrations vary in dependence from other genetic factors. Though the external appearance of brown and dun colored pigeons is very similar, the quantitative analysis of the melanins clearly separates the two genotypes. Thus, eumelanin and pheomelanin can sometimes be confused phenotypically as in brown and dun. This study concludes that from the optical impression, it would seem that depressing effects of dilute on eumelanin contents mainly affects the black form of the pigment.
There are for possibilities for dilute. D+ - Full color (not diluted), dP - Pale, d - Dilute, and dex - Extreme Dilute
The dilution happens where the pigments on the feathers that absorbs the light to show the color. Under the microscope, the pigment of an intense color like black and its dilute dun are exactly the same color (black) except that in the dilute there are smaller and fewer pigment granules, which creates appearance of another color -- diluted black, known as dun.
Reduced - r
 Reduced
is different gene than diluted and because it is color in demand by
many breeders, the quality of reduced birds are improved in
many pigeon breeds. The reduced gene mutation was discovered in 1951
by Carl Graefe. He saw it in his rollers and recognized the difference.
This
was
the beginning of the reduced gene as we know it today. Mr. Graefe,
curious with this new color, proceeded to cross and backcross this
color gene and perhaps because he was very knowledgeable breeder
in genetics, we now have reduced genes in many pigeon breeds. It
did not take him long to discover that it was a sex-linked dilution gene
and
recessive
to the wild-type. He discovered that reduced will influence every
color. "Pastel" is a word that best describes the influence
of the reduced gene. Most breeders tend to prefer reduced mated
with spread black. From this mating you would get reduced
on black and it would look like a silver bird with the wing shield
feathers
laced with a black edge. The
reduced pictures seen in this section are called rosy
necks. Notice the beautiful rosy necks and reduced bar and body
colors in
these
birds.
Thanks
to Dal M. Stone for putting this color into Birmingham Rollers.
According to Mr. Stone, the genetic make-up of these birds come
from the combination
of smoky, reduced, gimple factors, even they closely mimic the
appearance of dominant opal. Reduced and dominant opal
are
two
distinct
mutations located on different chromosomes. The unimproved version
of the rosy neck bird would show pinkish neck and pink bars and
would make it easier for us the see the tail band.
Reduced
is different gene than diluted and because it is color in demand by
many breeders, the quality of reduced birds are improved in
many pigeon breeds. The reduced gene mutation was discovered in 1951
by Carl Graefe. He saw it in his rollers and recognized the difference.
This
was
the beginning of the reduced gene as we know it today. Mr. Graefe,
curious with this new color, proceeded to cross and backcross this
color gene and perhaps because he was very knowledgeable breeder
in genetics, we now have reduced genes in many pigeon breeds. It
did not take him long to discover that it was a sex-linked dilution gene
and
recessive
to the wild-type. He discovered that reduced will influence every
color. "Pastel" is a word that best describes the influence
of the reduced gene. Most breeders tend to prefer reduced mated
with spread black. From this mating you would get reduced
on black and it would look like a silver bird with the wing shield
feathers
laced with a black edge. The
reduced pictures seen in this section are called rosy
necks. Notice the beautiful rosy necks and reduced bar and body
colors in
these
birds.
Thanks
to Dal M. Stone for putting this color into Birmingham Rollers.
According to Mr. Stone, the genetic make-up of these birds come
from the combination
of smoky, reduced, gimple factors, even they closely mimic the
appearance of dominant opal. Reduced and dominant opal
are
two
distinct
mutations located on different chromosomes. The unimproved version
of the rosy neck bird would show pinkish neck and pink bars and
would make it easier for us the see the tail band.
 Reduced is a single sex-linked
recessive gene but there is some hereditary problems associated with
hens laying poor quality
eggs which can affect hatchability. Unlike dilution where the pigment
production cut by half, reduced seems to cut it by about a quarter.
Reduced color changes a wild-type (blue bar) bird to gimpel opal
(rosy neck) where it changes a dark red/green neck to whitish color.
Reduced birds have a tendency to change quite a bit in the first
molt and it might take a reduced bird a couple of molts to reach
its true color. Dilute birds and reduced birds are short-downed
and almost naked when they were first born but the short-down in
reduced
is usually nowhere near the shortness produced by dilute.
Reduced is a single sex-linked
recessive gene but there is some hereditary problems associated with
hens laying poor quality
eggs which can affect hatchability. Unlike dilution where the pigment
production cut by half, reduced seems to cut it by about a quarter.
Reduced color changes a wild-type (blue bar) bird to gimpel opal
(rosy neck) where it changes a dark red/green neck to whitish color.
Reduced birds have a tendency to change quite a bit in the first
molt and it might take a reduced bird a couple of molts to reach
its true color. Dilute birds and reduced birds are short-downed
and almost naked when they were first born but the short-down in
reduced
is usually nowhere near the shortness produced by dilute.
The three Color Reduction possibilities are: R+ - Nonreduced or Full Color, r - Reduced, rru - Rubella
Just like the dilute, the reduced is also a single sex-linked recessive gene. If you mate a reduced cock to reduced hen, all the young would be reduced. If you mate a reduced cock to any color hen, all the hens would be reduced and all the cocks would be a color-carrying reduced. If you mate any color cock to reduced hen, then all the young cocks would be a color-carrying reduced, and all the hens would be father’s color not carrying the reduced gene.
Phenotype vs Genotype
Recall from the previous pages that genotype is the term used to describe the genetic makeup, as distinguished from the physical appearance, of an organism. The genotype determines the hereditary potentials and limitations of an individual. Among organisms that reproduce sexually, an individual's genotype comprises the entire complex of genes inherited from both parents. Sexual reproduction guarantees that each individual has a unique genotype. Phenotype is the observable physical or biochemical characteristics of an organism, as determined by both genetic makeup and environmental influences. In other words, phenotype is the expression (the actual trait that we see) of a specific trait, such as "ash-red" color in pigeons. Phenotypes result from the expression of an organism's genes as well as the influence of environmental factors and the interactions between the two.
In the examples below, we will not cover all the sex-linked and all the autosomal mutations but instead only look at genotype and phenotype of two birds and well only evaluate the sex linked b locus alleles reduced, dilute and autosomal recessive red mutations.
BA -
ash-red, B+ -
blue/black, b - brown
R+ -
Nonreduced or Full Color, r - Reduced, rru -
Rubella
D+ -
Full color (not diluted), dP -
Pale, d - Dilute, and dex -
Extreme Dilute
E+ -
Full Color non recessive red, e -
recessive red
GENOTYPE
|
PHENOTYPE
|
|||
Sex
Chromosome |
Autosomal |
This
bird is a cock bird since he has two functional
sex chromosomes.
He is heterozygous wild-type (+) Blue/black split for brown (b).
This bird also carries recessive red gene (e) but
cannot express
it in hererozygous state. He is non-reduced
or full color (wild-type) but because the dilute locus carries
two copies of the same gene, this bird
is actually diluted from blue
to dun.
| ||
b locus
|
Reduced
|
Dilute
|
Recessive
red
|
|
+
|
R+
|
d
|
e
|
|
b
|
R+
|
d
|
-
|
|
GENOTYPE
|
PHENOTYPE
|
|||
Sex
Chromosome |
Autosomal |
This
bird is a hen since she has 1 functional sex chromosome and 1
alleles for each sex-linked mutations. She is an ash-red (BA),
but because she carries two copies of the recessive red gene
(e//e), she
will present herself as a recessive
red
instead
of ash-red.
She has the wild-type gene on the reduce locus, but the dilute
locus carries a copy of the dilute gene (d), so this bird will
actually present herself as a recessive
yellow.
| ||
b locus
|
Reduced
|
Dilute
|
Recessive
red
|
|
BA
|
R+
|
d
|
e
|
|
- |
- |
- |
e |
|