Lethal Genes In Pigeons
By Arif Mümtaz
March
2012
Before we get into lethal genes, let’s get familiar with the wild-type genes. In genetics, we assume there is a wild-type gene or genes for each trait in all the chromosomes of any given species. It is imperative to understand that phenotype of the wild-type is the result of all the genes acting together to produce what we observe in a wild-type pigeon. Every single gene that makes the wild-type must be present and function properly for us to see all the characteristics of a wild-type phenotype. We compare the rest of the mutants against the wild-type genes. The genetic symbol for every gene in wild-type is the plus sign (+), representing everything pertaining to a wild-type pigeon.
 For
instance, in pigeons "Bar Pattern" is considered to be part
of the wild-type phenotype. Therefore, other alterations of the bar pattern,
like
"Check," "T-pattern Check"
and "Barless," are considered as mutations. In other words,
there is a wild-type gene at the location of every mutant gene. In
the case
of
pattern when
the mutant for Check is present the bird is a Check. Or when the
mutant for T-pattern Check is present the bird is a T-pattern Check.
Similarly, when all
other genes at any other loci are wild-type but the bird has two
copies (homozygous) of
Barless genes, then the phenotype of the bird will be Blue Barless
because Barless happens to be recessive allele of the pattern series.
However,
when the bird is homozygous for whatever wild-type happens to be
at the pattern locus, and also wild-type
at any other loci that can impact pattern, the bird will be a Bar.
Examples of other mutants that can impact pattern are mutations like
Spread, Recessive White, Recessive Red, and Albino. From the mating
results,
we discover and assign symbols if the mutant is dominant, recessive,
or co-dominant to that wild-type phenotype.
For
instance, in pigeons "Bar Pattern" is considered to be part
of the wild-type phenotype. Therefore, other alterations of the bar pattern,
like
"Check," "T-pattern Check"
and "Barless," are considered as mutations. In other words,
there is a wild-type gene at the location of every mutant gene. In
the case
of
pattern when
the mutant for Check is present the bird is a Check. Or when the
mutant for T-pattern Check is present the bird is a T-pattern Check.
Similarly, when all
other genes at any other loci are wild-type but the bird has two
copies (homozygous) of
Barless genes, then the phenotype of the bird will be Blue Barless
because Barless happens to be recessive allele of the pattern series.
However,
when the bird is homozygous for whatever wild-type happens to be
at the pattern locus, and also wild-type
at any other loci that can impact pattern, the bird will be a Bar.
Examples of other mutants that can impact pattern are mutations like
Spread, Recessive White, Recessive Red, and Albino. From the mating
results,
we discover and assign symbols if the mutant is dominant, recessive,
or co-dominant to that wild-type phenotype.
From the genetic science point of view, all wild-type genes that all together contribute to making wild-type phenotype are important and necessary for the survival of the species; whether the gene is coding for carrying oxygen properly from the lungs to cells, or it is the wing pattern of the bird. When we compare the mutant genes to the wild-type genes, we will also discover that the mutation under study can be beneficial, neutral, or lethal to the individual bearing the mutant genes. Not all mutations affect the organism because there is a certain amount of redundancy in the genetic information. If a mutation is "translated" from DNA into the protein that makes up the organism's structure, it may be in a non-functional part of the protein and thus have no detectable effect. This is known as a "neutral" mutation, and they tend to accumulate gradually as time passes. Some mutations do affect genes that control protein production or functional parts of protein, and most of these are lethal to the organism. Thus, when an essential gene is mutated, it can result in a lethal phenotype. Therefore, if a mutation destroys an important function of an unlucky individual then it can be lethal.
The definition of a lethal gene is any gene that results in the premature death of the organism, where dominant lethal genes kill in heterozygous state (carrying a single copy of the lethal gene), and recessive lethal genes kill only in homozygous state (carrying double copies of the lethal gene). In other words, recessive lethals in homozygous state happens when two alleles of the same gene which enter a zygote from the two parents are identical, such genes cause deviation from the normal development, and the individual is unable to survive. In addition, the recessive lethal genes in heterozygous state may reduce the viability of the individual carrying the mutation, but when lethal genes occur in homozygous state they produce the lethal effects. Dominant homozygous conditions have immediate lethal effect, while recessive lethal conditions show delayed effects and the gene may be transmitted to next generation. A lethal recessive allele is more likely to be maintained in a population if it is somehow beneficial in the heterozygous condition, or if it is not expressed until late in life. Rarely, mutations are detrimental to health; if the mutation is so severe, the individual dies before birth. More than likely we won’t see the dominant lethal genes that cause death in the early embryonic stage, because these individuals eliminate themselves from the gene pool and never have a chance to pass the gene to their offspring. These are called the "negative" mutations; a mutation that severely impairs the body's defense system against bacterial infection, for instance, would fall into this category. Most lethal genes are going to kill the individual at some point during the embryo stage, shortly after hatching, or birth. However, sometimes death caused by lethal genes is not immediate; it may even take years, depending on the gene. If death is caused just before the reproductive stage, they are called "sub-lethal" genes. The lethal genes topic would be way too long to cover, but let’s understand that other than dominant and recessive lethal genes there are also conditional lethal genes, semi-lethal or sub-lethal genes, and synthetic lethal genes.
 Let’s look at the
common lethal genes found in humans: Cystic Fibrosis, Tay Sachs, PKU
- Phenylketonuria, Sickle-Cell Anemia, Hemochromatosis, Neurofibromatosis
(NF), Huntington’s Disease, Hemophilia, etc. Let’s
take Sickle-Cell Anemia as an example: Sickle-Cell Anemia is an
abnormality
of hemoglobin, the molecule that carries oxygen into our blood. Hemoglobin
is contained within red blood cells. A mutation in the gene that
makes hemoglobin would be lethal if that mutation produced
a
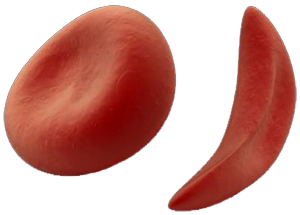
form of hemoglobin that could not carry oxygen. Human red blood cells,
which carry oxygen, are in the shape of turgid biconcave discs.
However, when a person suffers from sickle-cell disease, the cells
become sickle shaped, which reduces their oxygen carrying capacity.
This causes severe anemia, strokes and lung problems. Sickle-Cell
Anemia is an autosomal nearly recessive genetic blood disorder, characterized
by red blood cells of an abnormal, rigid, sickle shape. However,
there is an irony with this lethal gene. We know that some lethal genes
do
enough damage in some way to reduce viability but not enough to
kill every time. In fact, depending on the environment or the geography
in which the species exists, a lethal gene can prove to be beneficial
and kept
in the gene pool by a process called natural selection. If
an individual is heterozygous for Sickle-Cell Anemia, it enables that
person to withstand
malaria infections better than the individuals having wild-type
genes at that locus (specific physical location of a gene or DNA sequence
on a chromosome).
Let’s look at the
common lethal genes found in humans: Cystic Fibrosis, Tay Sachs, PKU
- Phenylketonuria, Sickle-Cell Anemia, Hemochromatosis, Neurofibromatosis
(NF), Huntington’s Disease, Hemophilia, etc. Let’s
take Sickle-Cell Anemia as an example: Sickle-Cell Anemia is an
abnormality
of hemoglobin, the molecule that carries oxygen into our blood. Hemoglobin
is contained within red blood cells. A mutation in the gene that
makes hemoglobin would be lethal if that mutation produced
a
form of hemoglobin that could not carry oxygen. Human red blood cells,
which carry oxygen, are in the shape of turgid biconcave discs.
However, when a person suffers from sickle-cell disease, the cells
become sickle shaped, which reduces their oxygen carrying capacity.
This causes severe anemia, strokes and lung problems. Sickle-Cell
Anemia is an autosomal nearly recessive genetic blood disorder, characterized
by red blood cells of an abnormal, rigid, sickle shape. However,
there is an irony with this lethal gene. We know that some lethal genes
do
enough damage in some way to reduce viability but not enough to
kill every time. In fact, depending on the environment or the geography
in which the species exists, a lethal gene can prove to be beneficial
and kept
in the gene pool by a process called natural selection. If
an individual is heterozygous for Sickle-Cell Anemia, it enables that
person to withstand
malaria infections better than the individuals having wild-type
genes at that locus (specific physical location of a gene or DNA sequence
on a chromosome).
Therefore, it shouldn’t be surprising to find a lot of people heterozygous for Sickle-Cell Anemia in populations where malaria is a common disease. Each year, malaria attacks about 400 million people, two to three million of whom succumb to the illness, and most malaria victims are children. So, we can look at having a single copy of Sickle-Cell Anemia mutation as an advantage, but 1/4 babies produced from the two hetero parents will be babies who is homozygous for Sickle-Cell Anemia mutation.
 Another
human genetic disease is called Cystic Fibrosis (CF) where the wild-type
gene for Cystic
Fibrosis makes a material
that degrades the mucus
in lungs. This degradation makes the mucus thin enough that the tiny hair
structures in lungs can sweep the mucus out. It is one of the most common
chronic lung diseases in children and young adults. It is a life-threatening
disorder. CF is caused by a mutation in the gene for the protein Cystic
Fibrosis Transmembrane Conductance Regulator
(CFTR). Although most people without CF have two working copies of the
CFTR gene, only one is needed to prevent Cystic Fibrosis. CF develops when
neither gene works normally and therefore has autosomal recessive inheritance.
If the individual is homozygous for a defective form of the Cystic Fibrosis
gene, the mucus build up leads to pneumonia and other lung infections,
and people with the problem seldom live more than a few years. CF is
a very
common mutation for people with ancestors from Europe.
Another
human genetic disease is called Cystic Fibrosis (CF) where the wild-type
gene for Cystic
Fibrosis makes a material
that degrades the mucus
in lungs. This degradation makes the mucus thin enough that the tiny hair
structures in lungs can sweep the mucus out. It is one of the most common
chronic lung diseases in children and young adults. It is a life-threatening
disorder. CF is caused by a mutation in the gene for the protein Cystic
Fibrosis Transmembrane Conductance Regulator
(CFTR). Although most people without CF have two working copies of the
CFTR gene, only one is needed to prevent Cystic Fibrosis. CF develops when
neither gene works normally and therefore has autosomal recessive inheritance.
If the individual is homozygous for a defective form of the Cystic Fibrosis
gene, the mucus build up leads to pneumonia and other lung infections,
and people with the problem seldom live more than a few years. CF is
a very
common mutation for people with ancestors from Europe.
The irony is that Cystic Fibrosis in hetero state is believed to have provided some survival benefit when the Black Plague swept Europe and killed two-thirds of the population. Some scientists even suggest that CF being hetero is of benefit in recovery from pneumonia as well. It is reasonable to argue that a mutation that gives the recipient an advantage over other individuals in a given population is less common. However, sometimes the mutation (Sickle-Cell Anemia and CF) improves the ability to survive a potentially deadly illness. The affected individual can then pass his/her genes to the next generation more efficiently than other individuals because they are more likely to reach reproductive age. This increases the chance that the modified gene will survive into the first generation and from there move into the following generations; thus, it is a "positive" mutation -- the mutation that may enable the mutant organism to withstand particular environmental stresses better than wild-type organisms or to reproduce more quickly.
 Now
that we are familiar with the definition of lethal genes and gave some
examples of lethal genes found in humans, let’s look at the common
lethal genes that we can identify in pigeons. One of them is called
dominant opal, which was first studied
and reported by W. F. Hollander and is proved to be an autosomal co-dominant
mutation -- genetic symbol
Od. We currently have no data to suggest
that in heterozygous state dominant opal pigeons have less viability
or have advantage against a certain disease,
but we know that dominant opal in the homozygous state is lethal. The
few suspected homozygous dominant opals that lived were near white,
showing a second distinct phenotype and therefore fit to the definition
of a co-dominant gene. As a side note, the other well defined co-dominant
genes found in pigeons are grizzle, indigo, and almond.
Now
that we are familiar with the definition of lethal genes and gave some
examples of lethal genes found in humans, let’s look at the common
lethal genes that we can identify in pigeons. One of them is called
dominant opal, which was first studied
and reported by W. F. Hollander and is proved to be an autosomal co-dominant
mutation -- genetic symbol
Od. We currently have no data to suggest
that in heterozygous state dominant opal pigeons have less viability
or have advantage against a certain disease,
but we know that dominant opal in the homozygous state is lethal. The
few suspected homozygous dominant opals that lived were near white,
showing a second distinct phenotype and therefore fit to the definition
of a co-dominant gene. As a side note, the other well defined co-dominant
genes found in pigeons are grizzle, indigo, and almond.
It seems that presumptive homozygotes have been reported with a variety of defects: whitish plumage color, ragged feathering, poor growth and small size, crippled feet, skin abnormalities causing behavioral problems, poor eye-sight, subcutaneous air puffs, and often severe tremor at a young age. Thus, dominant opal gene seems to have an impact on connective tissues. It is possible the accumulated connective tissue abnormalities are lethal. When two heterozygous dominant opal birds (Od//+) are mated, 1/4 will lack the Od gene (+//+), 2/4 will be dominant opals in hetero state (Od//+), and 1/4 will not hatch or usually die very young (Od//Od).
 The
other well-known and documented lethal gene is known as the web-lethal -- genetic
symbol wl. Webbed toes can be found fairly
commonly in many pigeon breeds but this occurrence certainly is not
evidence that pigeons evolved from a web-footed type of bird such as
gulls. Usually the webbing is between the inner and middle toes, or
even all three toes. Most of these webs are recessive, harmless and
autosomal mutations, and there seems to be relationship between web-feet
and feathered-feet. However, there has been some variation of the toe-webbing
where the webs do not always extend completely to the claws. This particular
webbing is a sex-linked lethal mutation and therefore it is near lethal
in the homozygous state for cock birds (wl//wl) and near lethal to
hens in the hemizygous state (wl//.). Hens only have one functional
sex chromosome and therefore are referred as hemizygous. Since hens
have no normal counterpart gene
on the W chromosome, a safety backup that compensates, any recessive
sex-linked mutation will present itself on hens.
The
other well-known and documented lethal gene is known as the web-lethal -- genetic
symbol wl. Webbed toes can be found fairly
commonly in many pigeon breeds but this occurrence certainly is not
evidence that pigeons evolved from a web-footed type of bird such as
gulls. Usually the webbing is between the inner and middle toes, or
even all three toes. Most of these webs are recessive, harmless and
autosomal mutations, and there seems to be relationship between web-feet
and feathered-feet. However, there has been some variation of the toe-webbing
where the webs do not always extend completely to the claws. This particular
webbing is a sex-linked lethal mutation and therefore it is near lethal
in the homozygous state for cock birds (wl//wl) and near lethal to
hens in the hemizygous state (wl//.). Hens only have one functional
sex chromosome and therefore are referred as hemizygous. Since hens
have no normal counterpart gene
on the W chromosome, a safety backup that compensates, any recessive
sex-linked mutation will present itself on hens.
Most of the times the death caused by web-lethal mutation will happen at hatching. Accrording to Hollander and Miller study on web-lethal mutation, majority of the abnormal (web-lethal) squabs either fail to hatch or fail to live over 10 days. Hollander and Miller were able to raise three survivors with peculiar eyes, where the iris was irregular and pupil looked pear-shaped or rectangular but vision seemed normal. This study concludes that even though web-lethal mutation seems to be quite inferior to normal, and generally lethal (causing death before maturity), perhaps the survivors have superior residual heredity.
Robert J. Mangile is one of the few people and probably the first person who ever produced and reported raising homozygous web-lethal cock (wl//wl). Until Mangile raised such a bird, it was not thought possible. He raised about 30 offspring from matings with the potential to produce wl//wl cocks. Mangile performed autopsy on them and found that some birds displaying the web-lethal trait (webbed or fused toes) had testes -- indicating maleness. According to Mangile, cock birds heterozygous for web-lethal (wl//+) did not display web toes but hemizygous hens (wl//.) and homozygous cocks (wl//wl) did. "Web-lethal hens (wl/.) and web-lethal cocks (wl//wl) both display similar symptoms of morbidity, i.e., affected eyes and pupils, small size, tubular shaped body, toes that vary from almost normal looking to rear & inner toes fused along with outer and middle toes webbed (not fused) - both in varying degrees, some with only one foot showing fused toes and/or webbed toes and some with near normal pupils. The greater the expression of this characteristics the more apt the possessor is to being less vital and/or vigorous," says R. J. Mangile (private communication).
 There
is another lethal gene found in pigeons called achondroplasia,
also known as dwarfism, which is a skeletal disorder, characterized
by
failure
of normal conversion of cartilage into bone. In an article written
by W. F. Hollander called "Lethal Achondroplasia in The Pigeons",
dwarfism found in pigeons as a recessive sex-linked lethal condition, where
the
embryo never hatches to survive, hence known as the lethal gene. Although
W. F. Hollander admits that further scientific research needs to be done
for lethal achondroplasia in pigeons, it is apparent that it does exist
in pigeons. He assigned the symbols ac for
this mutation. A lot of breeders think they might have a dwarf pigeon
in their loft because they have
extremely
short
legs
or
very
small
feet
or stubby
toes. Some birds are also a lot smaller than the rest, and the breeder
may assume it is caused by dwarfism. However, Hollander’s report
on achondroplasia (dwarfism) in pigeons is lethal, and the babies die in
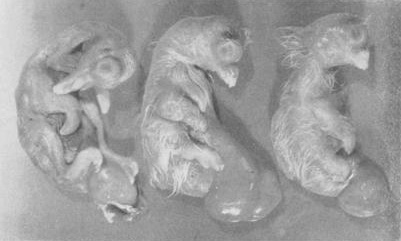
the egg before seeing the daylight. According to Hollander, the embryos
demonstrated on the right picture were found to be uniquely abnormal. None
of the
three embryos succeeded in breaking the eggshell; in fact, there
was no indication that even an attempt was made to break the
shell. Left: a normally proportioned embryo: center and right,
achondroplastic embryos,
male
and female.
There
is another lethal gene found in pigeons called achondroplasia,
also known as dwarfism, which is a skeletal disorder, characterized
by
failure
of normal conversion of cartilage into bone. In an article written
by W. F. Hollander called "Lethal Achondroplasia in The Pigeons",
dwarfism found in pigeons as a recessive sex-linked lethal condition, where
the
embryo never hatches to survive, hence known as the lethal gene. Although
W. F. Hollander admits that further scientific research needs to be done
for lethal achondroplasia in pigeons, it is apparent that it does exist
in pigeons. He assigned the symbols ac for
this mutation. A lot of breeders think they might have a dwarf pigeon
in their loft because they have
extremely
short
legs
or
very
small
feet
or stubby
toes. Some birds are also a lot smaller than the rest, and the breeder
may assume it is caused by dwarfism. However, Hollander’s report
on achondroplasia (dwarfism) in pigeons is lethal, and the babies die in
the egg before seeing the daylight. According to Hollander, the embryos
demonstrated on the right picture were found to be uniquely abnormal. None
of the
three embryos succeeded in breaking the eggshell; in fact, there
was no indication that even an attempt was made to break the
shell. Left: a normally proportioned embryo: center and right,
achondroplastic embryos,
male
and female.
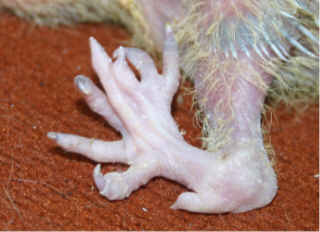
Polydactyly or polydactylism, which is a congenital physical anomaly found in some species having supernumerary fingers or toes, comes from the Ancient Greek polus, meaning "many," and daktulos, meaning "finger.” This mutation in pigeons was first reported on the Journal of Heredity in 1916, and was found in a Racing Homer from Australia. It was later studied by Hollander and Levi in 1939 and 1940. They reported this mutation as simple recessive gene and assigned the genetic symbol py. Sublethal polydactyly included blindness, the eyes, however, showed no evident structural abnormality. According to their study, a variable syndrome including polydactyly of feet and wings, shortened legs, undershot beak, short nestling down, and defective plumage was found in a family of Silver King pigeons. However, out of 25 specimens, none survived to maturity.
 Because
there seemed to be a lot of variations between the each individual
with the polydactyly mutation, Hollander and Levi decided to classify
the
polydactyls into four grades or groups. Group I had no extra thumb or only
on one wing; single extra hind toe, generally more or less joined with
other toe. Group II had a single extra thumb on both wings; single extra
hind toe on each foot, usually not joined with other toes. Group III had
single, usually large, extra thumb on each wing; two extra toes or claws
on one or both of the feet. Group IV had more than one extra thumb on each
wing; two or more extra toes or claws on one or both feet; feet distorted. According
to Hollander and Levi's
study on polydactyly mutation, since none of the 25 polydactyls survived
to maturity, in spite of their
efforts
to maintain them, they considered the py factor as sub-lethal mutation.
The immediate cause of death has not been found, and most of the specimens
seemed to
be thriving the day before death. The respiratory trouble noted in the
first polydactyl was also present in most of the others which lived to
the second week or beyond, and might have caused death by suffocation,
but ordinarily this ailment is not serious, and none of the specimens showed
internal lesions severe enough to be fatal. The polydactyls in group IV
(extreme) were so abnormal that they were unable to hatch.
Because
there seemed to be a lot of variations between the each individual
with the polydactyly mutation, Hollander and Levi decided to classify
the
polydactyls into four grades or groups. Group I had no extra thumb or only
on one wing; single extra hind toe, generally more or less joined with
other toe. Group II had a single extra thumb on both wings; single extra
hind toe on each foot, usually not joined with other toes. Group III had
single, usually large, extra thumb on each wing; two extra toes or claws
on one or both of the feet. Group IV had more than one extra thumb on each
wing; two or more extra toes or claws on one or both feet; feet distorted. According
to Hollander and Levi's
study on polydactyly mutation, since none of the 25 polydactyls survived
to maturity, in spite of their
efforts
to maintain them, they considered the py factor as sub-lethal mutation.
The immediate cause of death has not been found, and most of the specimens
seemed to
be thriving the day before death. The respiratory trouble noted in the
first polydactyl was also present in most of the others which lived to
the second week or beyond, and might have caused death by suffocation,
but ordinarily this ailment is not serious, and none of the specimens showed
internal lesions severe enough to be fatal. The polydactyls in group IV
(extreme) were so abnormal that they were unable to hatch.
Out of all the squabs they raised during this study, they were able to keep one female alive for 157 days. This bird, without the help of Hollander and Levi, would have died at much younger age under ordinary conditions, because her undershot beak made it difficult for her to pick up grain. They hand-fed her for some time after weaning, and if the feed and water had not been placed in the most obvious positions, she would have died a lot sooner. It was noted that her eyes appeared to be normal but they did not focus forward properly, and the mental development seemed poor. The bird could not fly, and walked awkwardly. Hollander and Levi concluded that this mutation, whether extreme or slight, leads to early death for individuals in homozygous state.
Although in humans it is a misconception to think that there is a twin gene and it is inherited, it is not totally accurate. There is a gene that causes hyper ovulation, which increases the tendency to release multiple eggs during ovulation, increasing the chances of conceiving fraternal twins. So, in families where the women have a gene for hyper ovulation, genetics would sufficiently explain an increased presence of fraternal twins. Some pigeon eggs have twins in them, but they are simply not able to hatch.
 I
can’t
explain the genetic reasons for why or how pigeons would have twins,
but it certainly leads to death of both twins. If it is caused by
some gene
and
it is inherited, perhaps we could add that to our list of lethal
genes found in pigeons. According
to a study done by Hollander and Levi about double yolk egg twins,
it was inferred
that crowding was a major factor
in preventing hatching. They have examined approximately three
dozen double-yolk eggs in this study and reported that all such eggs
were
very large; generally a normal-sized egg accompanied each in the
clutch, though occasionally both eggs of the clutch possessed two
yolks. Hollander
and Levi found that a few were infertile, but the majority showed
two fairly normal separate embryos. Death occurred usually during
the first
week of incubation. In one case an embryo died at the end of
the first week, but the other survived to term (17 days) and was decidedly
over-sized.
In addition, five large eggs developed both twins to hatching
stage.
Several broke the shell, but none actually emerged. All were
dead when placed in formalin. It was found that each embryo had its own
yolk-sac,
and was in no way attached to its twin.
I
can’t
explain the genetic reasons for why or how pigeons would have twins,
but it certainly leads to death of both twins. If it is caused by
some gene
and
it is inherited, perhaps we could add that to our list of lethal
genes found in pigeons. According
to a study done by Hollander and Levi about double yolk egg twins,
it was inferred
that crowding was a major factor
in preventing hatching. They have examined approximately three
dozen double-yolk eggs in this study and reported that all such eggs
were
very large; generally a normal-sized egg accompanied each in the
clutch, though occasionally both eggs of the clutch possessed two
yolks. Hollander
and Levi found that a few were infertile, but the majority showed
two fairly normal separate embryos. Death occurred usually during
the first
week of incubation. In one case an embryo died at the end of
the first week, but the other survived to term (17 days) and was decidedly
over-sized.
In addition, five large eggs developed both twins to hatching
stage.
Several broke the shell, but none actually emerged. All were
dead when placed in formalin. It was found that each embryo had its own
yolk-sac,
and was in no way attached to its twin.
Several other genes found in pigeons where the pigment production is much lower than normal, such as almond, reduced, dilute, lemon, albino, and the red phased recessive opal, also cause obvious health problems. These genes cause reduced life spans and poorer hatchability compared to other genes. Therefore, it is reasonable to consider all of these as weak semi-lethal mutants where in all of these cases it is clear that they are not any place close to fully lethal. For example, it is not recommended to mate two almonds together to make homozygous almond cocks. Almond cock birds in homozygous state usually die in the nest or, should they survive, will often have degrees of blindness or a clumsy character. However, several people including Carl Graefe reported that homozygous almond cocks do not have eye problems and presumably survived just fine provided that such birds are loaded up with darkeners. In fact, if they are loaded with enough darkeners (sooty, dirty, smoky, etc.) they were not even bald at hatching. Heterozygous almond cocks do not seem to have any health issues. In addition, with almond we have the curious situation where the hemizygous almond hens have normal viability even though hens do not have a balancing wild-type gene to help viability. Therefore, why homozygous almond cock birds have health problems but hemizygous hens don’t still remains a mystery to pigeon genetic science. According to Dr. Cryberg, there is another puzzle without any good answers about vitality of some mutations. “In the case of almond, reduced, dilute, lemon, albino, and the red phased recessive opal the pigment production is much lower than normal. There are parts of the brain that depend on some pigment intermediates for proper functioning. So perhaps some of the problem with these lighteners is those areas of the brain simply do not have enough of those intermediates? Yet, that does not quite make sense either as albino is not lethal,” says Dr. Cryberg (private communication).
 According to Dr. Cryberg,
although it might seem extreme and not justified, it is reasonable
to argue that every mutant we have is semi-lethal
as if
that were not the case the mutant would be common in wild pigeons. “In
all these cases it is likely that other genes are having some impact on
how viable the birds are with the result that in some cases the bird may
be so near normal in health, fertility and vigor that you cannot tell them
apart from wild-type while in other cases the same gene may result in dead
embryos or early deaths or abnormal eye structures,” says Dr.
Cryberg (private communication).
According to Dr. Cryberg,
although it might seem extreme and not justified, it is reasonable
to argue that every mutant we have is semi-lethal
as if
that were not the case the mutant would be common in wild pigeons. “In
all these cases it is likely that other genes are having some impact on
how viable the birds are with the result that in some cases the bird may
be so near normal in health, fertility and vigor that you cannot tell them
apart from wild-type while in other cases the same gene may result in dead
embryos or early deaths or abnormal eye structures,” says Dr.
Cryberg (private communication).
Lethal mutations, as the term implies, lead to the death of the individual. Death does not have to occur immediately; it could happen during the embryonic stage or may take several months or even years. But if the expected longevity of an individual is significantly reduced, the mutation is considered lethal. Evolution by natural selection will tend to remove lethal genes from the gene pool. However, a late-acting lethal gene may still be successful, as its effects will not harm the individual until it has had a chance to have some offspring.
Therefore, death from old age is a by-product of the accumulation in the gene pool of late-acting lethal genes, which have been allowed to slip through the net of natural selection simply because they are late-acting. Like many things in genetic science, the lethal genes topic is not fully understood and it is certainly not a black-and-white situation. Instead, lethal genes are simply a great big grey area that is hard to define. In any case, if a mutation results in lethality, then this is indicative that the affected gene has a fundamental function in the growth, development, and survival of an organism.
Lethal genes just allow changes or alter what goes on inside their possessor much like medicine where the small dose of a deadly toxin/chemical/medicine will make an individual sick, but a double dose has the potential to kill. For instance, let’s consider the effects of the wild-type genes at the recessive red loci. We know that wild-type gene makes some protein product that is part of what makes black pigment. If we have a bird that is heterozygous for recessive red (e//+), this bird can only produce one unit of functional protein as it only has one wild-type gene. On the other hand, if the bird has two copies of wild-type genes (+//+) at the recessive red loci, it will produce two units of functional protein as it has two wild-type genes. In both cases (e//+ and +//+), the phenotype of the bird will not have a dose response since recessive red is a pure recessive gene and can only alter the pigment production when an individual bird has two copies of the gene (e//e).
Now, let’s consider dominant opal. The dominant opal mutant in heterozygous state (Od//+) produces some protein that alters the chemistry that makes black pigment in some way. In heterozygous state, Od modifies the color of a wild-type plumage and make the bird appear as a typical dominant opal phenotype. However, in the homozygous state (Od//Od) the bird has a significantly different (near white) plumage. Thus we can clearly see the effects of single dose versus a double dose cause different phenotypic effects on the plumage color. In addition, double dose does more damage than one dose and it affects the vitality of the bird and not just the plumage color. The same vitality issues happens to the sex-linked almond mutation, where it has no known effects on hemizygous hens (St//.), but do have health issues in the homozygous state which can only happen to cock birds (St//St). In the homozygous state almond cock birds also show near white plumage compared to the heterozygous typical almond phenotype.
The more curious case exists in the sex-linked web-lethal mutation. It seems the cock birds carrying a single copy of the web-lethal (wl//+) gene don’t show any signs of abnormal feet or any known health problems. This means that the wild-type gene at the web-lethal locus produces something very important in allowing development of normal feet. We know that hens are hemizygous in their sex-chromosome and don’t have any allelic counterpart. However, although this idea is not fully tested, it seems hemizygous web-lethal hens (wl//.) survive better than homozygous web-lethal cocks (wl//wl) where breeding data shows homozygous cocks die more often as embryos. If this wild-type gene at the web-lethal locus is so essential to survive, why does web-lethal mutation show in the phenotype of hens, but not become as lethal for them? Even though breeding data suggest that homozygous cock birds die more often and only few of them were able to survive, we don’t have any complete statistical analysis of how many of each sex fail to hatch -- but we do know that many web-lethals of both sexes die before they hatch. Further research is needed to solve these mysteries of different lethal genes interacting with the wild-type genes and showing various vitality results depending on the presence or absence of other mutations.
If you have a closely related family of birds and experience more eggs never hatching than normal, you could have a recessive lethal gene of some sort in the gene pool of your family of birds. There are many genetic and epigenetic reasons why a chick never has a chance to hatch, but the intent of this article was not to cover all the remaining known and unknown reasons of unsuccessful hatching of the embryo. At some point of your pigeon breeding program, you might encounter the lethal mutations mentioned above or perhaps new mutations that no one has ever reported. Live stock breeding programs are aimed at fixing certain traits and removing others. Periodically, an unwelcomed trait may surface. When this happens, the breeder should ask if the mutation is desirable or undesirable. More than likely, the new mutation will be recessive because if it was dominant, it would have expressed itself prior. Undesirable dominant mutations are very easy to clean out of the gene pool because they express themselves with a single copy of the mutated gene. However, most of the time, the mutation will be recessive and will be very hard to take out of the gene pool, since it can be carried over a number of generations, and not present itself until two carriers of the mutated gene are mated. Even though eliminating recessive mutations from the gene pool can be a very difficult process and may not be done with 100% accuracy, it may be possible to further reduce the spread of these genes. Nature is designed so that mutations cannot be prevented, but controlled by either natural selection in the wild, or by artificial selection in breeding lofts.
Acknowledgments:
I would like to gratefully acknowledge the many valuable suggestions and corrections made by Dr. Richard Cryberg, who has also influenced me throughout this article. I would also like to give special thanks to Robert J. Mangile for sharing his extensive experience with the web-lethal mutation and for providing critical comments while writing this article.
References:
1. Anderson, W. W.
1969. Selection in Experimental Populations. I: Lethal Genes. Genetics,
62, 653–672.
2. Cryberg, R. PhD.
Private communication.
3. Donald F. Poulson. (2008) Lethal Gene. In
AccessScience, ©McGraw-Hill Companies. Retrieved March
12, 2012, from http://www.accessscience.com
4. Eaton, O. N. (1937). A Summary
of Lethal Characters In Animals and Man. Journal of Heredity. 28 (9)
: 320-326.
5. Hollander, W. F. (1945). Lethal Achondroplasia In The Pigeons.
Journal of Heredity Volume 36, Issue 10 Pp. 297-300.
6. Hollander, W. F. (1983).
Origins And Excursions In Pigeon Genetics: A Compilation. Burrton, Kan.: The
Ink Spot.
7. Hollander, W. F. and Levi, W. M. (Jul.,
1940). Twins and Late Embryonic Monstrosities in Pigeons The Auk Vol. 57,
No. 3 , pp. 326-329 Published
by: University of California Press.
8. Hollander,
W. F. and Levi, W. M. (1942). Polydactyl, A Sub-Lethal Character In The
Pigeon. Journal of Heredity Volume 33.
9.
Hollander, W. F. and Miller, W. J. (Oct 1982). A New Sex-Linked Mutation,
Web-Lethal from Racing
Homers. American
Racing Pigeon News.
10.
Instituto Gulbenkian de Ciencia (2011, April 28). Mystery Solved: How
Sickle Hemoglobin
Protects Against Malaria. ScienceDaily. Retrieved March
20, 2012, from http://www.sciencedaily.com /releases/2011/04/110428123931.htm
11. Levi, W. M. (1963). The pigeon (2d ed.). Sumter, S.C.: Levi
Pub. Co.
12. Mangile, R. J. Private communication.
13. Quinn, Joe W. (1971). The Pigeon
Breder’s Notebook.
QB&Q Publishing.
Copyright March, 2012 by Arif Mümtaz.