
9. X-Pigeons |
||||
 When
studying pigeon genetics, one of the most important concepts we
need to comprehend is wild-type.
In order to study the mutant traits, we choose the most common phenotype
from a population of organisms that represents the rest of that population.
It is just a standard phenotype which we test against other mutations.
The actual expression of those genes found in wild-type’s genotype
produces a set of observable characteristics in the wild-type’s
phenotype. The designation of wild-type is based upon a numerical
representation or estimation of the standard in a population. Although
living things seldom express the absolute perfection outlined in
a standard, they can be compared to one.
When
studying pigeon genetics, one of the most important concepts we
need to comprehend is wild-type.
In order to study the mutant traits, we choose the most common phenotype
from a population of organisms that represents the rest of that population.
It is just a standard phenotype which we test against other mutations.
The actual expression of those genes found in wild-type’s genotype
produces a set of observable characteristics in the wild-type’s
phenotype. The designation of wild-type is based upon a numerical
representation or estimation of the standard in a population. Although
living things seldom express the absolute perfection outlined in
a standard, they can be compared to one.
The ancestor to our domestic pigeons, the rock-pigeon (Columba livia), is considered to be the wild-type, the standard against which we judge everything. The Scientific name "Columba" is the Latin word for dove or pigeon; "livia" means blue or blue-gray color. Columba livia originated in Africa, Asia, Europe, and the Middle East. They arrived in North America with European settlers in the 17th-century for food and religious purposes. Because rock-doves in North America (and other continents) are from populations established by escaped domestic pigeons, we refer to them as feral pigeons. The term "wild" is reserved for pigeon populations that were never domesticated and still live in their native habitats - various open and semi-open environments such as cliffs and rock ledges which are used for roosting and breeding in the wild.
In pigeon genetics, the wild-type we use is commonly known as blue bar with a clean head - free from any head crest and has clean red pigmented legs - free from any leg feathers. Although we don’t have the exact measurements of all the phenotypic traits of the wild-type that traces back 150 years to use as a standard for everything, for color and a few other things it is very well defined. It does not have, for example, any heavy skin above the beak or around the eyes like the Barbs, Carrier Pigeons, and Kings do. They have a dark bluish-gray head, neck, and chest with glossy greenish, and reddish-purple iridescence along its neck and wing feathers. The neck and crop feathers are dark, with much iridescence. The adult female is almost identical to the male, but the iridescence on the neck is less intense and more restricted to the rear and sides, while that on the breast is often very obscure. Young birds show little lustre and are duller. The iris of the eye is red at the margin and yellow centrally, giving an orange appearance. The beak and the claws are very dark brown and black. For feather ornaments wild-type is defined as lack of feather ornaments (not frilled, fantailed, crested, has sideburns, longer neck feathers to cover its head completely like in Jacobins or Capuchines, etc.). The plumage is basically a light bluish gray; the remiges become black distally, and the ends of the rectrices also show black distally but in the form of a broad band with a faint bluish terminal edge. The tertiary wing feathers exhibit a black area in the vanes, as do secondary coverts and often the median coverts; these two rows of black areas form two black bars across the closed wing. The rump and under wing feathers are albescent or whitish terminally; the outer vanes of the outer rectrices are also albescent. Wild-type has 12 rigid tail feathers with a clearly distinguished dark area at the end of the tail known as the “sub-terminal tail band”. The name “albescent” was given by W. F. Hollander for those areas to distinguish from whiteness due to complete absence of melanin such as in white flights. Only the tail feathers on each side of the tail have the albescent strip on the outside, running from the base of the feather almost to the tail bar.
 We
also know that wild-type has oil (preen) gland above the base of their
tail, although not all domesticated pigeon
breeds have oil glands. This gland secretes oil when the pigeon grooms
itself, thus spreading this oil over its feathers. This oil contains
a compound that produces vitamin D when exposed to the sun’s
ultraviolet rays. So, as the pigeons grooms under an ultraviolet
light source, it’s actually mixing up a healthy batch of vitamin
D on its feathers. Reports
suggest that in general birds with extra tail feathers do lack the
gland above their tails; however, no one knows why. Our current understanding
of pigeon vision suggests that we don’t have the slightest
evidence of enhanced vision in wild-type compared to other mutants.
Although we don’t have any detailed explanation of why almond
mutation is causing eye problems, Carl Graefe reported highly pigmented
almonds did not seem to have many eye problems compared to almonds
with lack of the pigment in their feathers. In addition, we don’t
have any clue about the smelling ability of Columba livia or any
of the mutant breeds. A few
day-old wild-type squabs have a ring around their beak, just behind the
tip,
which lasts
about a week after the chick
was hatched.
The rest of the beak is sort of flesh colored and not highly pigmented.
However, if any of the darkener genes (mutations) are present in the
genotype the whole beak gets pigmented, and the beak ring does not show.
Up to five or seven days of age the legs of the wild-type should be flesh
colored and show no pigment at all. After that age, leg color will start
to very slowly turn red. If any of the the darkener genes are present
in their genotype, the
leg color turns brownish-blackish. Newly hatched squabs
also have distinct down colors, length and amount. Although we don’t
have any quantitative reports, there is a claim that cocks have a different
color down than hens do. However, there is a lot of variation in down
colors from near white to quite orange. Newly hatched wild-type squabs
considered to be normal down.
We
also know that wild-type has oil (preen) gland above the base of their
tail, although not all domesticated pigeon
breeds have oil glands. This gland secretes oil when the pigeon grooms
itself, thus spreading this oil over its feathers. This oil contains
a compound that produces vitamin D when exposed to the sun’s
ultraviolet rays. So, as the pigeons grooms under an ultraviolet
light source, it’s actually mixing up a healthy batch of vitamin
D on its feathers. Reports
suggest that in general birds with extra tail feathers do lack the
gland above their tails; however, no one knows why. Our current understanding
of pigeon vision suggests that we don’t have the slightest
evidence of enhanced vision in wild-type compared to other mutants.
Although we don’t have any detailed explanation of why almond
mutation is causing eye problems, Carl Graefe reported highly pigmented
almonds did not seem to have many eye problems compared to almonds
with lack of the pigment in their feathers. In addition, we don’t
have any clue about the smelling ability of Columba livia or any
of the mutant breeds. A few
day-old wild-type squabs have a ring around their beak, just behind the
tip,
which lasts
about a week after the chick
was hatched.
The rest of the beak is sort of flesh colored and not highly pigmented.
However, if any of the darkener genes (mutations) are present in the
genotype the whole beak gets pigmented, and the beak ring does not show.
Up to five or seven days of age the legs of the wild-type should be flesh
colored and show no pigment at all. After that age, leg color will start
to very slowly turn red. If any of the the darkener genes are present
in their genotype, the
leg color turns brownish-blackish. Newly hatched squabs
also have distinct down colors, length and amount. Although we don’t
have any quantitative reports, there is a claim that cocks have a different
color down than hens do. However, there is a lot of variation in down
colors from near white to quite orange. Newly hatched wild-type squabs
considered to be normal down.
We also have well defined behaviors of the wild-type. For voice, wild-type has the normal coo, and it is not equipped to sing like Trumpeters. For sexual behavior, wild-type is the normal behavior and not the exaggerated behavior we would see in birds like Spanish Thief Pouters. Flying pattern is also well defined where they fly in a range of height and distance from their home. They don’t have specialized mutations to roll, tumble, or hover like Sky Cutters, dive like Dewlaps, find home from long distances like Homers, or fly in other acrobatic patterns like Doneks and Kelebeks. They are not able to fly long hours like the high flying breeds averaging 10 hours or more flight time.
There are claims about Columba livia having magnetite in their beaks and brains for them to use Earth’s magnetic power to navigate. However, we have no clue which genes control this trait, and the standard amount of magnetite found in wild-type vs. homing pigeons is unknown. We currently don’t have any data about their blood type system or immune system genes to establish a standard either.
The genetic symbol for every gene in wild-type is the plus sign (+), representing everything pertaining to a wild-type pigeon. As you get familiar with distinct genes and their symbols you will notice that we don’t have a symbol for all the distinct traits. So far, we have done a fair job of characterizing traits that only involve one mutant gene which are mainly color mutations. There are some color mutants that involve several genes and we haven’t been able to isolate and characterize a single one of those mutants yet. The assignment of symbols does not occur until someone publishes data that is reproducible, sensible, and statically valid. We know that all traits are controlled by genes and we normally assign meaningful symbols to the mutated genes: al for albino, Od for dominant opal, b for brown C for checker, cr for crest, d for dilute, etc. In the case of wild-type, we assume that we do not know which genes control which traits, even though we can observe that wild-type has orange eyes, bluish body with two black bars and black sub-terminal tail band, etc. We are not allowed to give genotypical names to wild-type genes, since it is our standard. Therefore, we assign the wild-type symbol (+) to every gene in every locus.
 It
is imperative to understand that phenotype of the wild-type is the
result of all the genes acting together to produce what we observe
in a wild-type Columba livia. Every single gene that makes the wild-type
must be present and function properly for us to see all the characteristics
of a wild-type phenotype. Thus, no particular gene found in wild-type’s
genotype is special; they are all equally important and necessary.
It
is imperative to understand that phenotype of the wild-type is the
result of all the genes acting together to produce what we observe
in a wild-type Columba livia. Every single gene that makes the wild-type
must be present and function properly for us to see all the characteristics
of a wild-type phenotype. Thus, no particular gene found in wild-type’s
genotype is special; they are all equally important and necessary.
If any of the wild-type genes are broken (mutated) the whole bird is not going to function in a wild state as well as with a wild-type gene. In a mutated pigeon, the phenotypic difference may be small and may not even be visible to naked eye, but it will be real. It is not necessary to identify each of thousands of genes but only to know that the affect from the entire genome package all together produces a wild-type phenotype. It should then be obvious to observe which genes are affected to cause the mutations on an otherwise wild-type phenotypes. This way, the descriptions can be reduced to deviations from the standard. The mutant gene responsible for a deviation becomes the center of attention and a standard of reference minimizes the chance of mistakes in interpreting the results of breeding tests. Wild-type helps us understand the connection between presence or absence of a genetic trait in an individual.
We are assuming there is a wild-type gene at every locus on every chromosome in our standard Columba livia and any change affecting the function of any gene is called an allele at that location. These changes can be recessive, dominant or co-dominant and might be tightly or loosely linked to another locus. The phenotypic trait we observe may be and usually is dependent on more than one gene, or series of genes. We know that the mutated gene located at the dilute locus causes ash-yellow plumage if the wild-type gene at the ash-red locus is also mutated. When multiple mutated genes are present in a genotype, an interaction is apparently taking place and we get an ash-yellow plumage. Similarly, all the wild-type genes must be present and interact together to produce the wild-type phenotype. By adding a single mutant gene, e.g., dilution (d) to a female's genotype, the bird's phenotype is diluted in color (some fanciers call this phenotype "silver"). We can often identify the affect of a single mutant gene on a wild-type plumage - as well as on other mutants we've identified, e.g., dilute-brown (two mutants).
A pigeon that has wild-type genes in every loci of every chromosome is nearly impossible to get, as we don’t even have the sequencing of pigeon genome on hand to compare and identify which sequence codes for wild-type in any given locus. Then the question should arise how do we know if the mutation we are testing is actually tested against the true wild-type? From a practical standpoint, we do not even have a half decent source of wild-type to use as standards, so we do the best we can to find a bird that is very close to wild-type. In addition, in order to test a mutation, the wild-type bird we are testing against does not need to have wild-type gene in every single locus. Hollander pointed out repeatedly that we need to use a bird that is reasonably wild-type for the traits we are testing. Suppose we are testing grouse (feathered feet) against wild-type to see if it is dominant, recessive, or co-dominant. We can literally use any pigeon of any color (doesn’t have to be a blue bar) as long as the bird we are testing has clean legs like the wild-type and does not carry grouse mutant in its genotype (heterozygous for grouse). The same goes for beak crest, side burns, shell crest, etc. Therefore, we can use any bird that is wild-type for the particular mutation we are testing as long the bird does not carry that mutant gene in heterozygous state. It does not generally matter at all if the bird is far from wild-type in traits that are not under study. Now, if we are testing a color mutation, then we have to use a blue bar. However, it doesn’t matter if the blue bar (wild-type) has muffs, crest, pearl eyes, or fantail, etc. In other words, there is no reason at all to avoid using an ash-red, spread Tumbler Pigeon with a colored rump, that has muffs and has a shell crest in a study of tail feather count.
You might be wondering why we need a standard. According to Richard Cryberg, PhD, standards are simply picked and defined so that everyone uses the same thing; so, we can talk to each other and make sense. When studying genetics we choose a standard because it allows two people to talk about genetics without wasting all of their time defining the standard they are using every time they talk to a new person. Rock Pigeons display many variations among their numbers scattered throughout the world. The argument rages on yet, as to whether the earliest forms were barred or checkered. Why did they choose the blue bar phenotype as the wild-type in pigeons? Could they have chosen blue check as the wild-type instead? Or could they have chosen ash-red barless or spread black or any other common phenotype as our standard? The answer is “yes”. So, why did they choose the blue bar and not the check? The answer is very simple: whoever first defines the standard in any given species, whichever phenotype he/she chooses becomes the wild-type for that particular species.
In the 13th edition of Systema Naturea, Linnaeus Tom I.2 (1789), the author Gmelin has become the first person to describe the phenotype of the Rock Pigeon (the ancestor of our domestic pigeons) and named it Columba livia. He described it as ash-grey with a white rump and a black bar on the tail and wings, (“cinerea, uropygio albo, alurum fascia, caudaeque apice nigricante”). Gmelin knew the Rock Pigeon was the ancestor to domesticated pigeons, and the domestication took place more than four thousand years before Gmelin described the wild-type. According to Gmelin, the most important difference between the domestica (domesticated pigeons) and the livia (wild-type blue-gray pigeon), was the double bar on the wings (“alarum fascia duplici”). Because Gmelin was the first person to describe the phenotype of Columba livia, the genotype became the wild-type in pigeons--the median against which everything is judged. The rock pigeon/dove is the overwhelming choice that gave rise to modern pigeons. Plumage varies widely across the world, but the white-rumped blue bar is the phenotype that was selected, and it became our standard.
“It is sometimes difficult to decide what is normal. In the absence of a known wild-type one can use logic to help decide. For example, in cattle, horns to defend against predators or just in dominance conflicts are more likely than the absence of horns. Or the preponderant frequency of occurrence of one of alternative characters in a population can decide the choice. For example, are brown or blue eyes normal in humans? Consider the blacks of Africa. Almost all have dark eyes. Similarly the Chinese have brown eyes. Already we have more than half the world’s population. Add the American Indians, East Indians, Polynesians, Melanesians, …etc. and the world peoples preponderantly have brown eyes. If all else fails, one can make an arbitrary choice” (Wilmer J. Miller, A Survey of Genetics).
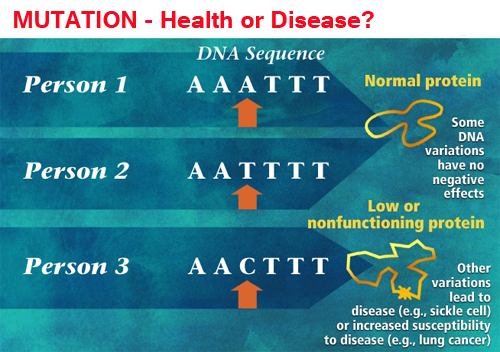 In
many organisms, alternatives (alleles) that are not a part of that
genotype of the wild-type are
considered as mutant alleles. Now and then, cells make
mistakes in copying their own DNA, inserting an incorrect base, or
even skipping a base, as the new strand is put
together. These mistakes are called mutations,
from the Latin word meaning "to
change". A mutation is a permanent
change in the DNA sequence of a gene. Sometimes, mutations in DNA can
cause
changes in the way a cell behaves. This is because genes contain
the instructions necessary for a cell to work properly. If some of
the
instructions to the cell are wrong, then the affect from changes
or errors in the cell alter the actions of the genes and produce a
different
effect. There
are two ways in which DNA can become mutated. Mutations can be
inherited if a parent has a mutation in
his or her DNA, and then the mutation is passed on to his or
her children. Mutations can also be acquired when environmental agents
damage DNA,
or when mistakes occur when a body cell copies its DNA prior
to
cell division. If a mutation takes place in somatic cells - any cell
of the body except sperm and egg cells, they will not be passed on
to the next generation. If, however, a mutation takes place in germ
cells (sperm or egg), then the mutations will be inherited by the offspring.
When the DNA strands are separated and copied, the altered base will
pair
with an incorrect
base and cause a mutation. In the example
showing on the left a "modified" T on Person 2 will
not make any effects and cause any problems. However, the incorrect
base from A to C in person 3 will cause major diseases.
In
many organisms, alternatives (alleles) that are not a part of that
genotype of the wild-type are
considered as mutant alleles. Now and then, cells make
mistakes in copying their own DNA, inserting an incorrect base, or
even skipping a base, as the new strand is put
together. These mistakes are called mutations,
from the Latin word meaning "to
change". A mutation is a permanent
change in the DNA sequence of a gene. Sometimes, mutations in DNA can
cause
changes in the way a cell behaves. This is because genes contain
the instructions necessary for a cell to work properly. If some of
the
instructions to the cell are wrong, then the affect from changes
or errors in the cell alter the actions of the genes and produce a
different
effect. There
are two ways in which DNA can become mutated. Mutations can be
inherited if a parent has a mutation in
his or her DNA, and then the mutation is passed on to his or
her children. Mutations can also be acquired when environmental agents
damage DNA,
or when mistakes occur when a body cell copies its DNA prior
to
cell division. If a mutation takes place in somatic cells - any cell
of the body except sperm and egg cells, they will not be passed on
to the next generation. If, however, a mutation takes place in germ
cells (sperm or egg), then the mutations will be inherited by the offspring.
When the DNA strands are separated and copied, the altered base will
pair
with an incorrect
base and cause a mutation. In the example
showing on the left a "modified" T on Person 2 will
not make any effects and cause any problems. However, the incorrect
base from A to C in person 3 will cause major diseases.
The genetic code has built-in redundancies, some mistakes might not have much effect on the protein made by the gene. In some cases, the error might be in the third base of a codon and still specify the same amino acid in the protein. In other cases, it may be elsewhere in the codon and specify a different amino acid. If the changed amino acid is not in a crucial part of the protein, then there may be no adverse effect. However, if the changed amino acid is in a crucial part of the protein, then the protein may be defective and not work as well or at all; this type of change can lead to disease. Other types of mutations in DNA can occur when small segments of DNA break off the chromosome. These segments can get placed back at another spot in the chromosome and interrupt the normal flow of information. These types of mutations (deletion, duplication, inversion, insertion, and translocation) usually have severe consequences. As you can see from the picture below that there are lots of possibilities for mutations and in general they change the sequence of the DNA enough that the gene’s normal function is altered.
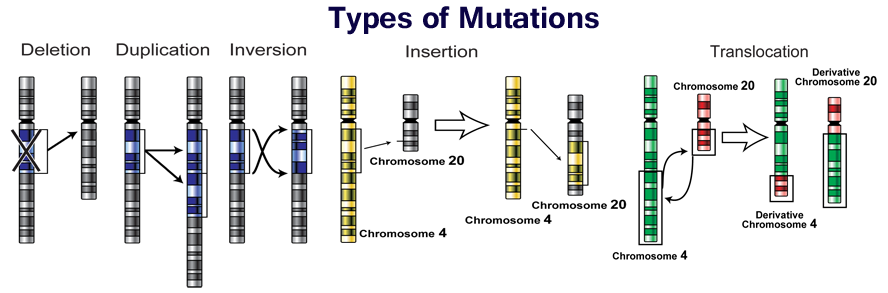
Errors in genetic information can cause diseases like cancer or birth defects such as Down syndrome. However, errors in genetic information are also the main cause that feeds evolution and thus create the great diversity of species and individuals. Most mutations are either neutral or harmful to survival and a small number of these changes will benefit survival. Most mutations that have any phenotypic effect are deleterious. Mutations that result in amino acid substitutions can change the shape of a protein, potentially changing or eliminating its function. This can lead to inadequacies in biochemical pathways or interfere with the process of development. Organisms are sufficiently integrated that most random changes will not produce a fitness benefit. Only a very small percentage of mutations are beneficial.
Mutation creates new alleles and each new allele enters the gene pool as a single copy amongst many. Most mutated alleles are lost from the gene pool, even the beneficial ones. The organism carrying them either fails to reproduce, or reproduces but does not pass on that particular allele. A mutant's fate is shared with the genetic background it appears in. A new allele will initially be linked to other loci in its genetic background, even loci on other chromosomes. If the allele increases in frequency in the population, initially it will be paired with other alleles at that locus -- the new allele will primarily be carried in individuals heterozygous for that locus. The chance of it being paired with itself is low until it reaches intermediate frequency. If the allele is recessive, its effect won't be seen in any individual until a homozygote is formed. The eventual fate of the allele depends on whether it is neutral, deleterious or beneficial.
"Remember, evolution is amazing -- but it isn’t perfect. Just about every adaptation is a compromise of sorts, an improvement in some circumstances, a liability in others. A peacock’s brilliant tail feathers make him more attractive to females -- and attract more attention from predators. Human skeletal structure allows us to walk upright and gives us large skulls filled with big brains -- and the combination means an infant’s head can barely make it through its mother’s birth canal. When natural selection goes to work, it doesn’t favor adaptations that make a given plant or animal “better” -- just whatever it takes for it to increase the chances for survival in its current environment. And when there’s a sudden change in circumstances that threatens to wipe out a population -- a new infectious disease, a new predator, or a new ice age -- natural selection will make a beeline for any trait that improves the chances of survival" (Moalem Sharon, and Jonathan Prince, Survival of the sickest: a medical maverick discovers why we need disease, p. 46).
Populations simply adapt to their current surroundings. They do not necessarily become better in any absolute sense over time. Organisms are not passive targets of their environment. In fact, each species modifies its own environment. At the least, organisms remove nutrients from and add waste to their surroundings. Often, waste products benefit other species. Animal dung is fertilizer for plants. Conversely, the oxygen we breathe is a waste product of plants. Species do not simply change to fit their environment; they modify their environment to suit them as well. Alternately, when the environment changes, species can migrate to suitable climes or seek out microenvironments to which they are adapted.
 As much as birds try to
adapt to their current environment, today the main threat posed to
birds comes from man, his destructive
tendencies and his manipulation of the environment. The life of birds
has changed enormously since man spread widely around the planet, and
began to develop and despoil it. Man is the primary force threatening
the natural world. Habitat destruction, hunting, introduction of alien
species and pollution combine to threaten almost 1000 species of birds
world-wide.
As much as birds try to
adapt to their current environment, today the main threat posed to
birds comes from man, his destructive
tendencies and his manipulation of the environment. The life of birds
has changed enormously since man spread widely around the planet, and
began to develop and despoil it. Man is the primary force threatening
the natural world. Habitat destruction, hunting, introduction of alien
species and pollution combine to threaten almost 1000 species of birds
world-wide.
Perhaps the best known bird extinction caused by man is the passenger pigeon (Ectopistes migratorius), a North American species. There was a population of an estimated two billion passenger pigeons in colonial days, when huge overflying flocks would darkened the sky. But even these phenomenal numbers could not guarantee the bird's existence. European colonists cut down the beech forests that provided food for the pigeons, and slaughtered the birds for food. The primary factor emerged when pigeon meat was commercialized as a cheap food for slaves and the poor in the 19th century, resulting in hunting on a massive scale. The last wild pigeon was shot by a boy in 1900; Martha, thought to be the world's last Passenger Pigeon, died on September 1, 1914, at the Cincinnati Zoo.
The Passenger Pigeon was larger than a Mourning Dove and had a body size similar to a large Rock Pigeon. The average weight of these pigeons was 340-400 grams (12-14 oz) and, length was 42 cm (16.5 in) in males and 38 cm (15 in) in females. The Passenger Pigeon had a bluish gray head and rump, slate gray back, and a wine red breast. The male had black streaks on the scapulars and wing coverts and patches of pinkish iridescence at the sides of the neck changed in color to a shining metallic bronze, green, and purple at the back of the neck in various lights. Female and immature birds were similar marked but with duller gray on the back, a lighter rose breast and much less iridescent necks. The tail was extremely long at 20–23 cm (8–9 in) and gray to blackish with a white edge. During summer, Passenger Pigeons lived in forest habitats throughout North America east of the Rocky Mountains from eastern and central Canada to the northeast United States. In the winters, they migrated to the southern U.S. and occasionally to Mexico and Cuba. However, the Passenger Pigeon's movements were not only seasonal, as with other birds, they would amass in whatever location was most productive and suitable for breeding.
When
natural population tends to increase in size, the individuals of the population
compete more for food, and other limited resources. Individuals of a population
differ from one another in the details of shared traits which have heritable
basis. These adaptive forms of traits make the bearers more competitive,
so those forms tend to become more common over generations. According
to Darwin,
this mechanism is called the Selection
of Traits.
In order for existence, survival and reproduction do not come down
to pure chance. Darwin
realized that if an animal has some trait that helps it to with stand
the environment better or to breed more successfully, it will leave
more offspring behind than others. On average,
the trait will become more common in the following generations. Darwin
thought random genetic mutations occur within an organism's genetic
code, and these beneficial
mutations are passed on to the next generation. Over time, beneficial
mutations accumulate and become an entirely different organism. 
Natural selection acts to preserve and accumulate minor advantageous genetic mutations. Natural selection is the only mechanism of adaptive evolution; it is defined as differential reproductive success of pre-existing classes of genetic variants in the gene pool. How does it work? Suppose a member of a species developed a functional advantage (it grew wings and learned to fly). Its offspring would inherit that advantage and pass it on to their offspring. The inferior (disadvantaged) members of the same species would gradually die out, leaving only the superior (advantaged) members of the species. Natural selection is the preservation of a functional advantage that enables a species to compete better in the wild. The ultimate causation, the broadcast, macro question facing Charles Darwin (picture on the right), was the evolutionary question: Why have factors in the environment pushed the birds this particular evolutionary direction? Why did this strange system evolve in reaction to evolutionary pressures? The ultimate explanation for birds to develop this ability is to give them enormous mobility to find food, escape predators, and seek better climates. We can look at natural selection as the naturalistic equivalent to domestic pigeon breeding. Over the centuries, human breeders have produced dramatic changes in domestic pigeon populations by selecting individuals to breed. Breeders eliminate undesirable traits gradually over time. Similarly, natural selection eliminates inferior species gradually over time. As Darwin studied natural selection, he spent a great deal of time with pigeon breeders, learning their methods. He joined two London pigeon clubs and attended pigeon competitions, where he connected with fanciers from around the world. He found their work to be an analogy for evolution. Thus, one of the organisms he study was the fancy pigeon, which he found to be a fountain of variation.
Evolution, or change in the gene pool of a population over time, is the process by which modern organisms have descended from ancient organisms. In order to understand evolution, it is necessary to view populations as a collection of individuals, each harboring a different set of traits. A single organism is never typical of an entire population unless there is no variation within that population. Individual organisms do not evolve; they retain the same genes throughout their life. When a population is evolving, the ratio of different genetic types is changing -- each individual organism within a population does not change. The process of evolution can be summarized in three sentences: Genes mutate. Individuals are selected. Populations evolve. Evolution is a reactive system since the evolving life-forms develop to survive in an environment which they do not control. The merciless process of the evolution of the human over its four million year history produced the intelligent warrior/hunter/explorer. The human became brave, smart, tenacious and capable of surviving under terrible mental and physical conditions. However, it is important to understand that evolution is not a planned (engineered) process and therefore it makes no attempt to build the optimum individual. Evolution creates life-forms without inherent direction or goal and leaves everything to pure chances without regard to the safety or comfort of the individual. Although not by design, this process insures that all possible physical life-forms and all possible combinations of life-forms are tested for survivability. As a result, variants with particular traits become more, or less, common. A key process in evolution is speciation, in which a single ancestral species splits and diversifies into multiple new species. There are several modes through which this occurs. Ultimately, all living (and extinct) species are descended from a common ancestor via a long series of speciation events.
 The
term Darwinism is Darwin’s specific
view that evolution is driven mainly by evolution & natural selection.
This theory, based on evidence
is suggesting that humans evolved from a common ancestor of apes
and human by the fusion of two pairs of chromosomes that reduced
the
chromosome number from 48 to 46.
The
term Darwinism is Darwin’s specific
view that evolution is driven mainly by evolution & natural selection.
This theory, based on evidence
is suggesting that humans evolved from a common ancestor of apes
and human by the fusion of two pairs of chromosomes that reduced
the
chromosome number from 48 to 46.
"Man has 23 pairs of chromosomes, while a chimpanzee has 24. There is only one pair that is different and this last difference is very interesting. If you look closely at human chromosomes one of them has two centromeres merged into a single centromere very close to the center of the chromosome. This is very unusual. And if you look at the specific letter sequences of this human chromosome and compare it with the various chromosomes in a chimp you discover that if you hooked two of the chimp’s smaller chromosomes together by combining the centromeres, both of which happen to be right at the ends, the letter sequence would be for all practical purposes identical to that single human chromosome. This indicates that sometime after humans and chimps each went its own evolutionary direction a chromosome fusion happened in the human line" (Taken from an unpublished article by Richard Cryberg).
Scientists believe this common ancestor existed five to eight million years ago, shortly after the species divided. One of these ultimately evolved into gorillas and chimps, and the other evolved into early human ancestors, called hominids. The Great apes (Bonobos, Chimpanzees, Orangutans, and Gorillas) are our wildlife cousins. We share a common bloodline - up to 98 percent of our DNA. We see a reflection of ourselves in their faces, in their mannerisms and in their interactions with kin. They have been on Earth for millions of years and have developed complex brains and complex societies. In the case of the striking similarities between human and chimp genomes, by far the simplest explanation anybody has ever proposed is that humans and chimps share a common ancestor. Because nobody has ever produced compelling evidence that this explanation is wrong, it is the one that scientists tentatively accept.
In 1895, when Charles Darwin proposed to the world the theory of evolution, the world was repelled. Many called it Gospel of dirt and attacked Darwins personal integrity and his motives. People rejected Darwins theory not because the evidence was against him. Most of the people who assailed him did not even understand any of his arguments. They rejected him because they were sickened by the fact that the science dragged human beings down to the level of the other animal species and diminished mankind. The people of the era, who were propounding an idealistic utopian view of a perfectible mankind, feared and despised Darwins theories.
 Many of the views which
have been advanced [here] are highly speculative, and some no doubt
will prove erroneous; but I have in every case given the reasons
which have led me to one view rather than another
The great principle
of evolution stands up clear and firm, when these groups of facts
are considered in connection with others, such as the mutual affinities
of the members of the same group, their geographical succession.
It is incredible that all these facts should speak falsely. He who
is not content to look, like a savage, at the phenomena of nature
as disconnected cannot any longer believe that the man is descendent
from a hairy quadruped, furnished with a tail and pointed ears, probably
arboreal in its habits
The main conclusion arrived at in this work
will,
I regret to think, be highly distasteful to many persons
But we are not here
concerned with hopes and fears, only with the truth as far as our reason allows
us to discover it. I have given the evidence to the best of my ability (Charles
Darwin, The
Descent of Man, 1871, Chapter 21).
Many of the views which
have been advanced [here] are highly speculative, and some no doubt
will prove erroneous; but I have in every case given the reasons
which have led me to one view rather than another
The great principle
of evolution stands up clear and firm, when these groups of facts
are considered in connection with others, such as the mutual affinities
of the members of the same group, their geographical succession.
It is incredible that all these facts should speak falsely. He who
is not content to look, like a savage, at the phenomena of nature
as disconnected cannot any longer believe that the man is descendent
from a hairy quadruped, furnished with a tail and pointed ears, probably
arboreal in its habits
The main conclusion arrived at in this work
will,
I regret to think, be highly distasteful to many persons
But we are not here
concerned with hopes and fears, only with the truth as far as our reason allows
us to discover it. I have given the evidence to the best of my ability (Charles
Darwin, The
Descent of Man, 1871, Chapter 21).
Darwin realized it was very difficult for people to accept his conclusion as to the origin of man. "Every one who admits the principle of evolution must see that the mental powers of the higher animals, which are the same in kind with those of man, though so different in degree, are capable of advancement. Thus the interval between the mental powers of one of the higher apes and of a fish, or between those of an ant and scale-insect, is immense; yet their development does not offer any special difficulty, for with our domesticated animals the mental faculties are certainly variable, and the variations are inherited. No one doubts that their mental faculties are of the utmost importance to animals in a state of nature. Therefore the conditions are favorable for their development through natural selection. The same conclusion may be extended to man; the intellect must have been all-important to him, even at a very remote period, as enabling him to invent and use language, to make weapons, tools, traps, etc., whereby, with the aid of his social habits, he long ago became the most dominant of all living creatures."
Domesticated Pigeon Breeds From Rock Pigeon
How did we get so many different breeds of domestic pigeons from the rock-dove like Jacobins, Pouters, Barbs and Rollers, Bokhara Trumpeters? Would they have adapted to survive with the new traits in the nature, if these mutations happened in wild populations? Did humans play a role in creating so many different pigeon breeds? Humans selected specific pigeons to reproduce certain traits in the hope they will make offspring better in some aspect than the parents. Thus, we helped pigeons evolve by noticing and collecting changes (mutations) a lot faster than nature could because we selected them more rigorously. We forced them to choose mates while in nature related birds very seldom would have mated. It is important to notice that most mutations that we currently see in our domestic pigeon breeds would not have helped their survival at all in nature. If they did, we would be seeing every pigeon roll or tumble, or sing like Bokhara Trumpeters. However, some mutations, like checker pattern protect them from the damaging ultraviolet lights in hotter climates and growing muffs for ice pigeon to keep their body heat, were beneficial mutations, and therefore we see them in nature. Even if rolling or tumbling for instance would be beneficial to their survival, it would have taken nature half a million years to get that mutation where man has done these things with pigeons in few thousand years.
Robert J. Mangile compares the effects of humanity with the effects of nature: "Unlike keepers of domesticated animals, ornithologists (bird scientists) assign great significance to seemingly minor plumage (and behavioral) variations. However, the significance is that such variations in wild species may occur in specific regions and the selection of these variations, even within a species, is done by the forces of Nature; and not by Man. And over the millennium Nature forges a new species, while Man forges a new breed. But regardless of whether Man or Nature does the selecting, the end result will most likely be considered an advancement - by Man. Nature makes no judgment calls!"
For thousands of years, humans have acted as agents of genetic selection, by breeding offspring with desired traits. All our domesticated animals (dogs, horses, cattle) and food crops (wheat, corn) are the result. Pigeon breeding is a case of artificial selection. One form of a trait is favored over others under contrived, manipulated conditions – in an artificial environment. Darwin saw that breeding practices could be an easily understood model for natural selection, a favoring of some forms of a given trait over others in nature. Just as breeders are “selective agents” to promote reproduction of certain pigeons, agents of selection act on the range of variation in the wild. Swifter and better camouflaged pigeons are more likely to avoid predators like falcons and hawks and live long enough to reproduce, compared with not-so-swift or too-flashy pigeons. Darwin taught us that the gene selection, either in the wild or human-directed (animal breeding), builds fantastically different creatures. They either better fit the niche in the ecosystem or better to respond to some breeders desire for a particular trait. The highly-varied domestication of Columba livia was Darwin's simple beginning for his theories. Darwin bought and kept every known pigeon breeds in England. Darwin knew if these fancy birds would be wild, they would have been classed as distinct species. Pigeon breeding gave Darwin the most graphic example of how species originate. The pigeons played such an important part in Charles Darwin's work on genetics and inheritance when he wrote “The Origin of Species". Darwin's case for his theory of natural selection therein has its own simple beginning, and not one having to do with an ancient or exotic organism, but with the species Columba livia, the humble rock-pigeon.
 Darwin wrote: Great as
are the differences between the breeds of pigeon, I am fully convinced
that all are descended from the rock-pigeon, Columba livia. Pigeon breeds which are
so different from one another, provided Darwin with compelling evidence
that the potential for evolutionary change was, in fact, present
in a single species, Columba livia. The implication was that, if
such variation could be achieved with pigeons by means of breeding
choices over the course of human history, then, perhaps, it was not
unreasonable to imagine that the enormous variety of the biological
world could have been achieved by means of natural processes over
the vast expanse of geological time. A
pigeon breeder selected individual birds to reproduce in order to produce
a head crest.
Similarly,
nature “selects” individuals
better suited to surviving their local conditions, because those
who are not as well adapted die off when the ones with an advantageous
trait may survive, because of that trait.
Darwin wrote: Great as
are the differences between the breeds of pigeon, I am fully convinced
that all are descended from the rock-pigeon, Columba livia. Pigeon breeds which are
so different from one another, provided Darwin with compelling evidence
that the potential for evolutionary change was, in fact, present
in a single species, Columba livia. The implication was that, if
such variation could be achieved with pigeons by means of breeding
choices over the course of human history, then, perhaps, it was not
unreasonable to imagine that the enormous variety of the biological
world could have been achieved by means of natural processes over
the vast expanse of geological time. A
pigeon breeder selected individual birds to reproduce in order to produce
a head crest.
Similarly,
nature “selects” individuals
better suited to surviving their local conditions, because those
who are not as well adapted die off when the ones with an advantageous
trait may survive, because of that trait.
Given enough time, Darwin argued, mutation and natural selection might produce new types of body parts, from wings to beaks. Darwin thought any ancestral population could generate, over time, individuals quite unlike themselves. With more than four thousand years of breeding behind them, domestic pigeons are full of combinations deemed beautiful or useful, or both, by their breeders. According to Darwin domestication seems to induce variation, but domestication does not increase the rate of mutation; it does increase the probability of the production of homozygotes.
The varieties of pigeons we see today are from the odd mutations that man noticed and liked, then bred the trait into his flock to intensify it. While man was attempting to create a new breed from an odd mutation, which would normally not have survived in the wild, nature used natural selection to ensure that the fittest and the strongest survived. Mutation and natural selection are not random events. Those mutations that allow the organism to survive and leave more offspring are preferentially accumulated within the gene pool; those that don't are preferentially removed (along with the animal bearing them). Variation in the wild is a necessity that allows for adaptation, which in turn encourages survival. Darwin thought that we don’t see mutations like archangels fantails, pouters, jacobins, feather ornaments, and crest in the wild because they would not have survived in the wild on their own. Human played a big role for these mutations to be evolved and perfected over time, resulting in well over 300 recognized pigeon breeds today.
How do we prove that all the fancy pigeons came from Columba livia, the rock-dove? When Darwin was studying pigeons, and wrote “The Origin of Species”, he devoted the whole first chapter to the domestication of pigeons. He discovered that if the different fancy breeds are mated, the offspring would eventually lose their distinctive traits and resemble the rock-dove. Darwin called this phenomenon “reversion”. According to Darwin, there was no other reasonable explanation for the profusion of fancy pigeon. Many pigeon breeders, who have been around pigeons for long time experience a reversion, where two distinct pigeon breeds of different colors and patterns are bred, their offspring show the body type and the color of an ordinary rock dove (wild-type). It is imperative to understand that the word reversion does not mean that genetic makeup is lost when the offspring tends to resemble the wild ancestor. When two different breeds of pigeons crossed recessive mutations from wild-type in each breed used in the cross are hidden, but not lost. So, recessive mutations will not show when a cross is made between a homozygous recessive mutant and a homozygous dominant mutant, only recessives genes undergo reversion.
 Hollander explains atavism,
regression, reversion to ancestral type with a less refined language
and calls it “throwbacks”.
In page 23 of his book “Origins and Excursions in Pigeon
Genetics,” He
gives examples of few crosses and throwbacks.
Hollander explains atavism,
regression, reversion to ancestral type with a less refined language
and calls it “throwbacks”.
In page 23 of his book “Origins and Excursions in Pigeon
Genetics,” He
gives examples of few crosses and throwbacks.
According to Hollander, if we take a purebred Damascene cock and cross it with a barless Coburg Lark, all of their progeny will look practically like wild Blue Rocks making all the cherished characteristics of each parent disappear. Each parent had recessive genes in the homozygous state compared to wild-type but not against each other where Damascene is iced, while Lark is barless and dilute. So the hybrids will not show any of these characteristics, instead resemble the wild-type blue bar. However, the offspring will not lose ice, barless or dilute genes and when the F1’s are mated to each other they will produce variety of phenotypes that they carry in their gene pool. In another example, let’s assume we have a recessive red bird that has wild–type genes in every other loci. We also have a milky bird that has wild–type genes in every other loci. Both milky and recessive red are recessive mutations and they can only be expressed when they are homozygous (carrying two copies) of the same gene. When they are mated, every single F1 from this mating will be blue bars—reversion to wild-type phenotype even though they will still be carrying recessive red and milky in their genotype. Therefore, reversion does not mean reverting back to original wild-type and losing the mutations forever.
Hollander also points out that some crosses don’t give reversion in F1 but maybe later generations. The example he uses is when a Black Barb is crossed with White Fantail. He doubts that the F1 hybrids will show any resemblance of the wild-type but in F2 some young may revert all the way. Because the characteristics of the pure parents were not all recessive, recombination in F2 may eliminate them and produce throwbacks.
“Knowing the recessiveness or dominance of the various domestic characteristics and the composition of each breed or variety, we can predict quite reliably what to expect from most crosses. We can plan, we can dodge, we can make Mendel jump through hoops. The only trouble is, we do not yet know all about each domestic characteristics”. (Hollander, Origins and Excursions in Pigeon Genetics, p. 23).
Mutations happen all the time and they are naturally not preventable because the DNA’s built-in mechanism to proofread errors during copying does not always catch those errors. In genetics, a mutation is a change of the nucleotide sequence of the genome of an organism. Mutations normally result from unrepaired damage to DNA or to RNA genomes from errors in the process of replication, or from the insertion or deletion of segments of DNA by mobile genetic elements. Nature has developed proofreading techniques that eliminate most errors but cannot correct 100% of them because some are unavoidable due to the laws of physical chemistry.
So, if mutations happen when the sequence of DNA is changed from what we consider the wild-type gene, is it possible for these so-called mutated versions of the genes to reverse back to the original wild-type sequence? The answer is very rarely, but yes. Therefore, a mutation can occur in two directions: mutation from wild-type to mutant is called a “forward mutation”, and mutation from mutant to wild-type is called a “back mutation” or “reverse mutation”. The reverse mutation refers to undoing the base sequence change from the mutant gene back to its original state (wild-type – true version) resulting in regained gene functionality. Although, it is hard to imagine mutants undergoing a reverse mutation back to wild-type version, it seems reasonable for DNA replication to make a mistake in copying again and bring the mutated gene sequence back to the functional wild-type state. Reverse mutations are very rare but can happen as long as the mutation was due to a single base change. For deletion or insertion mutations the reverse mutation is going to be so rare there is little or no hope of ever seeing it happen.
According to Dr. Cryberg, in pigeons we know of one example where two recessive red LFCL Tumblers produced a blue offspring. The person who owned these birds owned only red and yellow LFCL Tumblers and no other colors. “Typically there are lots and lots of ways to mutate a gene so it no longer functions. But once you have such a mutation there are only a very few changes that will allow the gene to function again. So, there are much greater odds that wild type would mutate to recessive red (or any other recessive) than the odds that recessive red would revert back to wild type,” says Dr. Richard Cryberg.
Because the word “reversion” that Darwin once described gives the impression that the mutant gene is lost and cannot be passed on to the future progeny as in “reverse mutation”, it is important to distinguish “reverse mutation” from what is known as “reversion”. For instance, if we mate an otherwise wild-type recessive white bird with an otherwise wild-type recessive red, all the F1 will be wild-type (blue) carrying both recessive red, and recessive white in heterozygous state. When these F1’s are mated together, they will show both of these recessive traits in F2. Therefore, the word reversion does not mean that genetic makeup is lost or changed forever when the offspring tends to resemble the wild ancestor. When reversion happens, these recessive genes inherited from both parents will be recovered in the future generations. However, in the case of reverse mutation, the change in a mutant gene restores its ability to produce a functional protein and, therefore, will remain as a permanent change.
Rigorous Selection of Traits
Humans have been transforming pigeons into some of the most bazaar creature on the planet. Domestic pigeons have outrages colors, crests, beaks, feathered feet, etc. Some of them even blow up like balloons looking nothing like their ancestors, while some of them competing for best in pigeon shows for their outstanding phenotype display. Pigeon breeds vary mostly by color, flying pattern, size, and feather shape. Some breeds, like the English Pouter or Capuchines barely resemble the typical shape of the traditional pigeon. The pigeon called the Jacobin, named after the hooded monks of France, is estimated to be a 500 year old breed. Scandaroons are estimated to be 2000 year old breed from Iraq. Frillbacks are the century old German breeds. Oriental Frills are ancient breeds from the Middle East showing shorter beak caused by genetic selection. But genetic manipulation found in domestic pigeons is far more than skin deep. Modenas are the angriest type and known as the avian Rottweiler. Some are breed for their aerial performance like Birmingham Rollers. They look like ordinary pigeons but they have been breed for their unusual way of flying. These domesticated breeds could not live in the wild on their own and more than likely will be assimilated or became extinct. In fact, Oriental Frills’ beak is so small, they can’t even feed their young - they need to be foster parented by pigeon fanciers. The Frillback Pigeon’s feathers will not help them stay warm and dry because they are not water resistant, and artfully flying Rollers can be easy target for bird of prey (BOP). Thus, pigeons are manipulated into extraordinary variety of shapes and sizes by humans via artificial selection of traits.
The pictures shown below are few examples of show type pigeon breeds that we have today. Some of them are so unique that most people don't even realize they're looking at pigeons. These mutants are all developed and reproduced with the help and careful selection of humans for more than four thousand years of breeding and domesticating the rock doves for variety of reasons. There are many, many breeds of domestic pigeons all with different appearances and behaviors. Almost any anatomical feature is different between the breeds; beak size and shape, feather coloration, feather orientation, some have feathered feet, some scaly feet, some webbed feet, some are very thin and tall, others fatter and larger, some have enlarged nasal skin appendages, some can expand their throats into a puff almost the size of their bodies, some have tail feathers which fan out like a peacock, almost all bones vary in dimension. Some have frill feathers or feathers covering their heads (hoods) like the Capuchines and Jacobins. Even behaviors change: some have odd coos, others have an odd flight pattern where they ‘tumble’ out of the sky. It is very important to understand that most of these mutations from the rock dove is independently inherited. If we want, we can make homing pigeons with crests or feathered feet. We can also put any color and pattern to any breed if we want to. Most of the traits are recessive to wild-type and therefore will take two or more generations to put in, but they are all possible to put in to any pigeon breed. In Genetics, it is therefore assumed that such traits are separate changes from the original rock-dove and majority of them inherited independently from each other.














