
9. X-Pigeons |
||||
 When
studying pigeon genetics, we need to be familiar with the
terms Genotype and Phenotype because
what we see on the outside (phenotype) does not necessarily determine
what that living thing actually has on the inside (genotype). Thus, to
achieve a modicum of breeding success, it is important that
pigeon breeders
understand the difference between - the reproductive potential
and the external appearance of an individual bird.
When
studying pigeon genetics, we need to be familiar with the
terms Genotype and Phenotype because
what we see on the outside (phenotype) does not necessarily determine
what that living thing actually has on the inside (genotype). Thus, to
achieve a modicum of breeding success, it is important that
pigeon breeders
understand the difference between - the reproductive potential
and the external appearance of an individual bird.
Genotype is the term used to describe the genetic makeup, as distinguished from the physical appearance, of an organism. Some genes are hidden, and we cannot see them by looking at the physical appearance. The genotype determines the hereditary potentials and limitations of an individual. Among organisms that reproduce sexually, an individual's genotype comprises the entire complex of genes inherited from both parents. Sexual reproduction guarantees that each individual has a unique genotype. Phenotype, on the other hand, is the observable physical or biochemical characteristics of an organism, as determined by both genetic makeup and environmental influences as well as the interactions between the two.
You may have heard about the Human Genome Project. Its goal was to find out the sequence of the 3 billion nucleotides pairs that make up a humans genome. A genome is all of an organisms genetic material. Unless you have an identical twin, you have a unique genome that determines all of your traits that can be seen, such as the color of your eyes. Other traits cannot be seen, such as the exact chemical makeup of your eyeball. Phenotype is the expression (the actual trait that we see) of a specific trait, such as the ash-red or recessive red in pigeons. A male pigeon, for example, can express itself as an ash-red bar in its phenotype. However, this particular bird could have lots of other hidden (recessive or masked) genes like recessive white, recessive red, brown, or any number of wild-type genes that together contribute to making blue/black in its genotype. We simply won’t know what this ash-red cock bird has in his genotype until we compare his mating results, or sequence his DNA.
Sex-linkage
As you continue to study pigeon
genetics, you
will be hearing about sex-linked genes, because it is
important to
understand this concept in order to predict the breeding results
of sex-linked genes. Each human has two chromosomes that determine
the
sex
and they are called X and Y chromosomes. In females, a dominant allele
on one of
the X sex chromosomes will mask a recessive allele on the other
X. In males, a recessive allele on the X sex chromosome will show
up
because there
is no matching allele on the Y. In sex-linked inheritance, alleles on sex
chromosomes are inherited in predictable patterns since males and
females have different sets of chromosomes, XX and XY. Typically
the X chromosome is longer and it bears a lot of genes not found
on the Y chromosome. Thus, most sex-linked genes are X-linked genes.
Sex linkage is the phenotypic expression of an allele related to
the chromosomal sex of the individual. 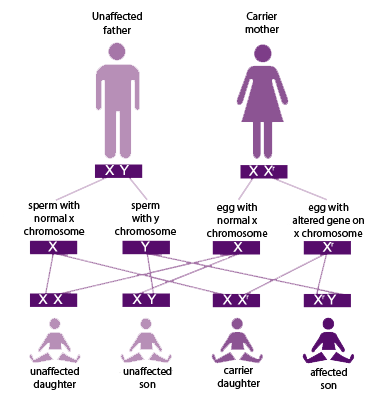
Let us look at sex-linked inheritance in humans to understand this concept. In humans, two well-known X-linked traits are hemophilia and red-green color blindness. Hemophilia is the failure to produce certain substances needed for proper blood-clotting, so a hemophiliac’s blood doesn’t clot, and (s)he could bleed to death from an injury that a normal person might not even notice. Hemophilia is much more common in males than females because hemophilia is located only in the X chromosome. Males are heterozygous (XY) - they only have one copy of the gene (X) in question - and therefore express the trait when they inherit one mutant allele. In contrast, a female must inherit two mutant XX alleles (homozygous) to have hemophilia, a less frequent event since the mutant allele is rare in the population. Females with only one affected X chromosome will be carriers.
Another example of sex-linked recessive inheritance is color blindness in humans. Color blindness is much more common in males than females because this mutation is also located on the X chromosome. Males have no alternative copy of the X chromosome to correct this problem, but females have a second X, a safety backup that compensates. If a female human has one copy of this mutant gene, she will just be a carrier but will have normal color vision. For females to have hemophilia or color blindness, they must inherit both affected X chromosomes from both of their parents. This reduces the chances of inheritance compared to male humans. A mother with one copy of the gene has a 50% chance of passing it to her children of both sexes, but her daughters will just be carriers of the gene unless their father has it, too. The picture on the right demonstrates the mating of an unaffected father and a carrier mother. If a male human is hemophiliac or color blind, he must have gotten this X gene from his mother, and if a female human is a carrier, she could have gotten the affected X gene from either of her parents. Thus, by definition hemophilia and color blindness are sex-linked recessive genes located on the X chromosome.
Pigeons have no external sex organs so it's often difficult to tell a male from a female. But if we know the pedigree (ancestry) of the father and the mother of the offspring, and if their phenotypic traits are sex-linked, we can tell the gender of the offspring by looking at its phenotype. In pigeons, ash-red, brown, almond series (faded / frosty / qualmond / sandy / hickory), reduced (rubella), and dilute (pale/lemon) are located in the sex-chromosome and therefore they are sex-linked. Recall that in pigeons cock birds have ZZ and hens have ZW, and as far as we know, there are no known genes located in the W. Therefore, hens have no backup copy for the sex-linked genes and during mating they can only donate the functional Z or non-functional W. Knowing this, we can easily predict the offspring of sex-linked mating results.
For instance, if you breed a hemizygous almond hen to a wild-type (blue/black) cock, all the cock birds from this mating will be heterozygous almonds. Because the hen must donate the only functional Z chromosome to make the offspring a cock bird (ZZ), and since almond is dominant to wild-type, all the male offspring from this mating will be almonds split for wild-type. All the hens from this mating will have a wild-type phenotype. Say we have a recessive red cock bird, but he is carrying a dilute (sex-linked) gene in one of his Z chromosomes. He cannot show dilute in his phenotype because dilute is a recessive gene and requires two copies in cock birds to show in their phenotype. Females, on the other hand, will show the dilute phenotype with a single copy since they only have one Z. Thus, if we breed a recessive red cock bird carrying dilute to a recessive red hen, 50% of the females from this mating will be recessive yellow. Recessive yellow is the diluted version of recessive red. Since all the recessive yellows we get from this mating will be hens, we conclude that dilute is a sex-linked gene. Let me give you another example. Ash-red mutation is dominant to wild-type. Thus, if we breed an ash-red hen to a wild-type cock bird, we expect all the cock birds from this mating to be ash-red and all the hens to be blue/black color. The sex-linked concept in pigeons will be explained in more detail in the next pages when we talk about the pigeon's plumage colors and pigments. Just try to remember that normally recessive genes need two copies to present themselves. However, in female pigeons we can observe all the sex-linked recessive genes, even if they are found singly in the genotype. Hopefully, you are now familiar with the sex-link inheritance of certain traits in pigeons.
Mitosis & Meiosis
"Normal" cells contain two full sets of chromosomes which are described as diploid ("diploos" is Greek for "double"). Our body cells are diploid - they contain two sets of chromosomes. In humans, a cell nucleus contains 23 pairs of chromosomes, or 46 individual chromosomes. Half of these chromosomes come from one parent and half come from the other parent. However, there is one kind of cell that is not diploid. Mature gametes, the sex cells (sperm and egg), have only one full set of chromosomes. Cells with only one set of chromosomes (gametes) are called haploid ("Haploeides" is Greek for "single").
When we talked about cell division before, we talked about how a mother cell divides and make daughter cells, and daughter cells divide and make their own daughter cells, etc. We call this division mitosis, often called "copy division" because the genetic material is copied. Mitosis provides additional cells for growth or replacement for dead or dying cells, and it is important to understand that in mitosis, the chromosome gene combinations remain unchanged unless the cell makes mistakes copying itself.
In order for our body to properly function, the body's cells must act cooperatively and function normally. Under normal circumstances, our cells behave consistently well and divide only when they are told, not straying far from their designated place of origin and carefully tracking their chromosomes. Even though our bodies generally do a good job at keeping things running smoothly, some ratio of mistakes is expected during cell replication process of mitosis. However, it is imperative to understand that the mistakes made during mitosis will not be inherited by the offspring, so any harmful effects will be limited to the individual who has abnormal cells. Such mistakes (mutations) are only passed to daughter cells during the process of mitosis and could cause some major problems for this individual or may not do any harm at all depending on the mistake the cell made during mitosis.
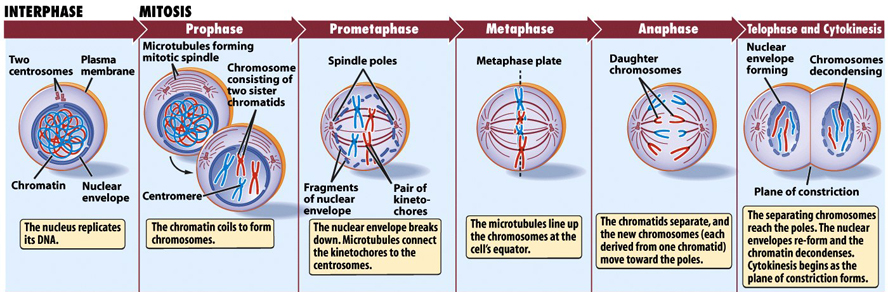
Cells come in all sizes and shapes. They perform thousands of different functions in the lifeforms of Earth. They work together to form the tissues of all living things. However, we know that cells do not live forever and they eventually die! Fortunately, they have the ability to reproduce first, through a process called cell division. It begins with a chemical signal to prepare for reproduction. The cell grows larger. It’s called the “G1 phase” of a cell’s life. After growth, the cell duplicates its DNA. This is called the “S Phase” of the cell cycle. The DNA is then checked by enzymes for mistakes and repaired if there are any. We talked about the error-checking mechanism on the previous page. This is the “G2 phase” of the cell cycle. When ready, cell division begins and the cell divides.
Mitosis begins when the nuclear membrane dissolves and the DNA thickens where chromosomes become visible, and disorganized (Prophase). Special fibers called spindle fibers form on each end of the cell and grow toward the DNA chromosomes. They attach to each chromosome of the homologous pair, resulting in a tug-of-war. The fibers then attach to centromeres on the chromosomes and pull them into a line in the middle of the cell (Metaphase). The centromere of each chromosome breaks down the middle and the chromosomes are pulled apart by the spindle fibers to opposite sides of the cell (Anaphase). Last is Telephase, where two new nuclei form. Nuclear membrane forms around both bundles of chromosomes at each pole. This creates two nuclei. Also, the chromosomes begin to unwind (decondense) until eventually they are nothing more than a dim coloring of chromatin with a few nucleoli. The cell then divides into two cells (Cytokinesis).
 All
of us started life as a single cell that divided again and again.
But that first
cell experienced something special. Cell replication within
the testes of males and ovaries of females is different form of
cell division and replication. We
call this division meiosis and it is often called "reduction
division" because the genetic material is reduced
by half. Meiosis is a two-part cell division process which occurs
in organisms that sexually reproduce.
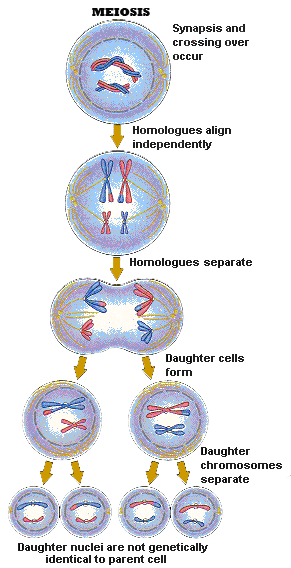
Meiosis produces gametes with one half the number of chromosomes
in the parent cell. Meiosis starts with a single diploid cell and when
meiosis is completed we have four haploid cells — cells with
a single set
of chromosomes. These haploid cells mature into sperms
or eggs. In some respects,
meiosis is very similar to the process of mitosis, yet it is also fundamentally
different. The
two stages of meiosis are meiosis I and meiosis II. At the end of
the meiotic process, four daughter cells are produced. Each of the
resulting daughter cells has one half of the number of chromosomes
in the parent cell.
All
of us started life as a single cell that divided again and again.
But that first
cell experienced something special. Cell replication within
the testes of males and ovaries of females is different form of
cell division and replication. We
call this division meiosis and it is often called "reduction
division" because the genetic material is reduced
by half. Meiosis is a two-part cell division process which occurs
in organisms that sexually reproduce.
Meiosis produces gametes with one half the number of chromosomes
in the parent cell. Meiosis starts with a single diploid cell and when
meiosis is completed we have four haploid cells — cells with
a single set
of chromosomes. These haploid cells mature into sperms
or eggs. In some respects,
meiosis is very similar to the process of mitosis, yet it is also fundamentally
different. The
two stages of meiosis are meiosis I and meiosis II. At the end of
the meiotic process, four daughter cells are produced. Each of the
resulting daughter cells has one half of the number of chromosomes
in the parent cell.
Thus, the main difference between mitosis and meiosis is the end result. In meiosis one diploid mother cell makes four unique haploid daughter cells instead of making two unchanged copies of diploid cells for growth. Meiosis only happens during the creation of sperm and egg cells, and it is extremely important for sexual reproduction so that each individual will have equal number of chromosomes form both parents. Meiosis produces four transposable unique haploid cells; it takes place only at certain times in an organism’s life cycle and is only involved in sexual reproduction. Mitosis produces genetically identical cells that result in diploid cells; it takes place throughout an organism’s lifetime and is only involved in asexual reproduction. In meiosis, each of the resulting chromosomes in the gamete cells is a unique mixture of maternal and paternal DNA, ensuring that offspring are genetically distinct from both parents. A gamete (from Ancient Greek means "husband" or "wife") is a mature sexual reproductive cell that unites with another gamete (sperm or egg) during fertilization (conception) in organisms that reproduce sexually. A zygote is a single cell created by the union of two sex cells (gametes). Life begins with the fertilization of an egg by a sperm, and a zygote is a new complete single cell united by these two gametes (sperm and egg).
Why are we reading about meiosis in pigeon genetics page? Well, the mistakes the DNA makes copying itself during meiosis, which creates sperms and eggs, can become a mutation and will be passed on to the next generations. If there is a mutation occurs during the meiosis, the mutation can be incorporated into a gamete. If that gamete is the one that eventually fuses with another gamete (i.e. if the mutated sperm fertilizes the egg), that mutation will be passed on to the offspring. As all the offspring's cells are the result of the first two gametes, all the organism's cells will have that mutation. Obviously this can have dire consequences for the offspring, if the mutation is harmful. For us the pigeon breeders, DNA making a mistake copying itself and creating new mutants is the reason why we have so many different colors and varieties of different pigeon breeds in the world. It certainly took thousands of years to accumulate all these mutant genes in our domestic pigeons, but mutations happen all the time; hence, it is imperative to understand how these mutations happen and how they are passed on to the next generations. Recall that if the mutation happens during mitosis (body cells) the mutant will not be passed on to the next generation. However, if the mutation happens during meiosis (sperm and egg cells) then the offspring will receive that mutation.
One might think meiosis is the only division responsible for reproduction, but is not entirely true. Many species, including some multicellular species, reproduce by mitosis and therefore they get the exact copies of the parents. This method of reproduction is called asexual reproduction the offspring from this reproductions are called clones because they are produced without sex and by using mitotic division. Some animals especially invertebrates use this mitotic way of reproduction, but plants are “masters” at cloning and that is why asexual reproduction is often called vegetative reproduction. Humans don’t use cloning method to reproduce but, once the zygote is formed, mitosis dominates and is responsible for making every nucleus in your body except the gametes (sperm and egg cells).
Now let’s talk about one of the most important parts of the meiosis. When an individual undergoes meiosis his sperm or her egg will not be a recreation of its father's sperm or mother's egg. Instead that individual will produce unique gametes containing a quarter of dad's genes and a quarter of mom's genes. That unique gamete will fuse with another (haploid) gamete and that fusion will create another unique zygote that will grow into a unique individual. Therefore, that new individual will be composed of the genetic materials from 1/4 of mom's mom, 1/4 of mom's dad, 1/4 of dad's mom, 1/4 of dad's dad. Each individual is unique but is also a mix. We are "half like" (1/2) each of our (two) parents, a "quarter like" 1/2) each of our (four) grandparents, an "eighth like" (1/8) each of our (eight) great grandparents, a "sixteenth like" (1/16) each of our (sixteen) great, great grandparents, etc. In humans, a cell nucleus contains 23 pairs of chromosomes, which are 46 individual chromosomes. For human gametes, with 23 pairs of chromosomes, the number of possible combinations of chromosomes a human can have from both of his parents is 223 or 8,388,608 alternative ways. In other words, each individual human could have been constructed 8,388,607 more ways and that could have made us look like a completely different person. The gametes will normally end up with 23 chromosomes, but the origin of any particular one will be randomly selected from paternal or maternal chromosomes. A pigeon has 40 pairs of chromosomes, which means pigeons have 80 individual chromosomes. The number of possibilities in pigeons is 240 or 1,099,511,627,776 possible combinations. One might think this is a very large number of possibilities. However, during gamete production, the paired chromosomes can exchange genetic material (crossover), increasing the number of possibilities mentioned above. This contributes to the genetic variability of progeny, a genetic descendant or offspring.
Crossover (Recombination)
Most of our cells are diploid with two sets of chromosomes. One set is from our mother and the other set is from our father. Each maternal chromosome has a corresponding paternal chromosome. These match pairs are called homologues chromosomes. During interphase chromosomes are duplicated where each chromosome consist two identical copies called sister chromatits. We know that chromosomes contain a very large number of genes. Those that are part of the same chromosome are said to be linked and to demonstrate linkage in genetic crosses. Because the chromosome, not the gene, is the unit of transmission during meiosis, linked genes are not free to undergo independent assortment. Instead, the alleles at all loci of one chromosome should, in theory, be transmitted as a unit during gamete formation. However, in many instances this does not occur. In meiosis homologues chromosomes stick together in pairs. The close association of homologous chromosomes allows segments of non-sister chromatits to trade places. This recombination of maternal and paternal genetic material is a key feature of meiosis and this mechanism is known as the crossover. During meiosis the DNA segments of matching chromosomes exchange small parts of each other between maternal and paternal homologues, and thus create new chromosomes that are not inherited solely from either parent. In other words, crossover occurs when two chromosomes, break and then reconnect but to the different end piece so that part of the first become part of the second and vice-versa. If they break at the same place or locus in the sequence of base pairs, the result is an exchange of genes, called genetic recombination. Genetically then, crossovers are new combinations, recombinations, of link genes. The crossover process is so exact that in the finished product there are exactly as many genes as existed in the starting chromosomes. The mechanism is very unique and perfect that even a gene can break in the middle for the swap and the replacement gene picks right up on the letter sequence so nothing is lost during crossover.
Let’s use a deck of cards analogy to explain crossover of genes between homologous chromosomes. Let’s say we have two full decks of cards: one deck is blue and the other deck is red. Let’s also assume these cards are not shuffled and their order remains constant. Let’s say these two decks of cards together represent a pair of chromosomes where the red deck is the maternal chromosome and the blue deck is the paternal chromosome. Each card in this deck is a gene; thus, the 10 of spades in the red deck has the corresponding 10 of spades in the blue deck, and the king of hearts is found in both decks, and so on. Both decks (homologous chromosomes) have exact number of cards (genes). If either the blue or the red deck is passed on to the next generation as a whole, we would know no genes were crossed over or exchanged. However, suppose the jack of diamonds has been swapped between the decks. The jack of diamonds of blue deck is now in the red deck, and the jack of diamonds of the red deck is in the blue deck. Hence, they have changed exactly the same number of genes. Now both decks have 51 of the same color cards and 1 different color in them. This tells us that crossover has happened. Easy, right? Well, that’s the simplest way to explain crossover of genes between homologous chromosomes.
When studying pigeon genetics, most people probably will not read or hear much about meiosis and mitosis but these concepts are very important to understand the crossover genes of chromosomes. Crossover is often overlooked by pigeon fanciers although it happens all the time during sexual reproduction. We simply cannot see or notice every single crossover occurrences because not all of them affect their phenotypes. Without the crossover, each gamete would get exact set of either paternal or maternal chromosomes without shuffling. In other words, without crossover, all alleles for those genes linked together on the same chromosome would be inherited together. When two or more genes reside on the same chromosome, whether sex chromosome or autosome, they are said to be linked. Linked genes do not independently assort during gamete formation. The crossover mechanism allows homologous non-sister chromosomes to exchange segments of varying length with each other and this allows linked genes to be separated and donated independently.
Let’s
take two sex-linked mutations found in pigeons, ash-red and dilute
for example. Suppose
we have a cock bird with a desired mutation (ash-red) located on
his paternal chromosome,
and we have
another desired mutation
(dilute) located on his maternal chromosome. We know this bird
presents itself as ash-red since ash-red is dominant to blue/black (wild-type),
and it is not diluted because in cock birds, the dilute mutation needs
two copies to make that bird diluted. So, when he makes
sperms during meiosis, he can either make sperms from his paternal
chromosome that has the ash-red at the b locus and wild-type at
dilute locus, or from his maternal chromosome that has the wild-type
at the b locus and dilute at the dilute locus.
Recall that he can only donate
one of his sex chromosomes to his offspring. Therefore, without
genes
crossing
over from one chromosome
to the other, all of his daughters will be either ash-red (non-diluted)
or dun (diluted blue/black). So, what about the possibilities of having
ash-yellow (diluted version of ash-red) or non-diluted blue/black daughters?
Crossing
over swaps some of the genes around
that wouldn’t typically be swapped and therefore it
could allow this cock bird to produce four possible phonotypic daughters
(ash-red, ash-yellow, dun, or blue/black), instead of only two unshuffled
chromosomes (ash-red or dun). Thus, because of crossover mechanism,
we are able to produce diluted
versions of all colors, ash-yellow, recessive yellow, dun, silver, khaki,
etc.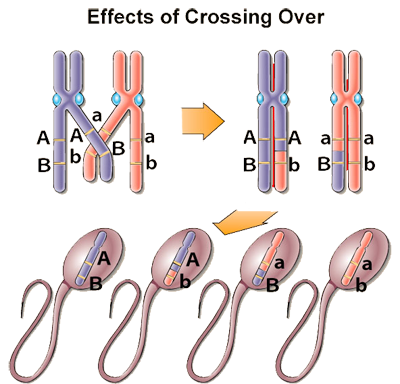
Take a look at the picture named "Effects of Crossing Over" on the right. What you are seeing in this picture is two homologous sex chromosomes. Homologous chromosomes are chromosome pairs of the same length, centromere position, and staining pattern, with genes for the same characteristics at corresponding loci. One homologous chromosome is inherited from the organism's mother; the other from the organism's father. In this picture, we see two sex chromosome of male human. Let’s say the purple chromosome is inherited from his father, and the pink chromosome is inherited from his mother.
During the 1st step (prophase I) of meiosis, when this individual starts producing sperms in his testis, homologues chromosomes come on top of each other before separating and this is when the swapping genetic material (crossover) between chromosomes occurs. If you look at the picture above, the one that shows meiosis, you will notice that during the 1st phase of meiosis each haploid chromosome makes copies of themselves and then form tight associations with each other. They then exchange DNA in the process of crossing over of genes. Notice that on the "Effects of Crossing Over" picture, before the crossover occurs this individual received dominant A and dominant B genes from his father (they are both on the paternal chromosome), and he received recessive a and recessive b genes from his mother (they are both on the maternal chromosome). Therefore, without the crossover he could have only created two unique combinations of chromosomes, which would have been AB, AB, or ab, ab; the exact copies of combinations he inherited from each of his parents. In other words, without crossing over, each haploid cell produced will inherit a chromosome with the same combination as was on one of the parental chromosomes. Thus, the arrangement of linked genes on the same chromosome is not altered in this case.
From the "Effects of Crossing Over" example, notice that the allele B of the paternal chromosome is crossed-over with allele b of the maternal chromosome. In this example, since the crossing over is only between two individual chromatids, we now have four different genetic combinations of sperms: AB, Ab, aB, and ab, where two sperms Ab and aB are added as the different possible combinations. Two will come from the chromatids that did not participate in crossing over and this will still have the parental combination of alleles, either A and B or a and b. The other two will be recombinant, with either A and b or a and B alleles. Thus, crossing over allows for reassortment of linked genes. In other words, crossover in this example created four differently combined sperms instead of the only two differently combined sperms. The only difference between the two situations is the statistical outcome.
Crossover rate can be affected by sex, age, temperature, proximity of allele to the centromere, inversion and many other factors. It also can vary from individual to individual a great deal. Every chromosome is expected to undergo at least one crossover during meiosis; most will have more than one crossover. There are hot spots where lots of crossovers happen. There are areas such as the centromere (cold spots) where crossovers are inhibited. The ratio of crossover between genes that are very closely located to centromere therefore is very, very low. We don’t fully understand the hot spots and cold spots but it is evidently due to epigenetics since hot spots are not the same in individuals of the same species. There is growing evidence that crossover rates in one individual may differ quite a bit from another individual even if both are the same sex. When two genes are located on the same chromosome, the chance of a crossover producing recombination between the genes is related to the distance between the two genes. Thus, the use of recombination frequencies has been used to develop linkage maps. However, it is important to note that recombination frequency tends to underestimate the distance between two linked genes. This is because as the two genes are located farther apart, the chance of double or more crossovers between them also increases.
 The
highest probability of crossover that could be seen between any two
genes for any species is no more than 50%. If crossover
was to
occur
at its maximum frequency (~50%) then the alleles would appear to
be independently assorting. Therefore, no
matter how far
apart the genes
are recombination cannot exceed 50%. This will be true even if the
actual probability of having a crossover between these two genes
is 99.99%.
It is perfectly possible to have genes where the probability of having
at least one crossover between those two genes for all practical
purposes is 100%.
The
highest probability of crossover that could be seen between any two
genes for any species is no more than 50%. If crossover
was to
occur
at its maximum frequency (~50%) then the alleles would appear to
be independently assorting. Therefore, no
matter how far
apart the genes
are recombination cannot exceed 50%. This will be true even if the
actual probability of having a crossover between these two genes
is 99.99%.
It is perfectly possible to have genes where the probability of having
at least one crossover between those two genes for all practical
purposes is 100%.
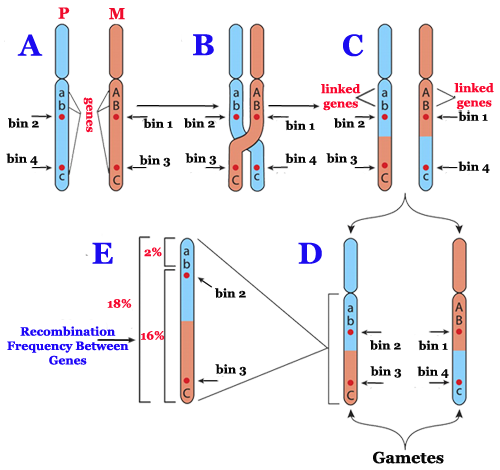
The Figure on the left is the Genetic Linkage and Recombination (A) Two homologous chromosomes: blue (paternal) and orange (maternal). Three genes with separate alleles and linkage disequilibrium "bins" noted (A,a; B,b; C,c; bins 1 to 4). (B) Crossing over during meiosis. (C) Two alleles and their linked bins (C,c; bins 3 and 4) have switched locations via recombination. Four additional alleles and their associated bins (A,a; B,b; bins 1 and 2) have not switched and are considered linked. (D) Recombined haploid chromosomes segregate separately during meiosis as gametes before fertilization. (E) Sample recombination frequencies between genes demonstrating higher rates of recombination for genes further apart.
It is important to understand that every single chromosome is expected to undergo at least one crossover during egg or sperm formation, it is therefore not a rare event at all. Crossovers are under genetic control so that the recombination of chromosomes does not happen too often. However, it is not unreasonable for a single chromosome to undergo several crossovers every single time. It is estimated that during meiosis in humans, there is an average of two to three crossovers for each pair of homologous chromosomes.
As we breed pigeons we need to be aware that the crossover mechanism can affect any pairing. The frequency of recombination between two locations (crossover) is depends on their distance between the genes involved. Genes close to one another have a lower frequency of crossover than do genes farther apart. In other words, we can say that longer the chromosome, and farther apart the genes from each other on the chromosome, more the chances of crossover. Thus genes located close together tend to be inherited together, which is called genetic linkage. Linked genes will be inherited in the same predicted ratios as would a single gene. In contrast, genes that are far apart are most likely to assort independently. Genetic linkage has let scientist calculate the physical distance between two genes. We don’t exactly know the distance between the loci of chromosomes. All we know are the crossover percentages and this tells us very little about the physical distance between the genes. The crossover % between the almond and brown locus is about 3 or 4%. The crossover % between reduced and dilute is speculated to be about 2 or 3% but no one has ever actually presented any data to support this number. The crossover rate between brown and dilute is about 45%. None of these numbers are well established. To get more accurate estimates would require raising a couple of thousand young.
The purpose of a crossover is to allow an equal amount of DNA material between pairs of chromosomes to be swapped from one chromosome to the other without changing overall sizes of each chromosome. The idea behind crossovers is that the new chromosome may be better than both of the parents if it takes the best characteristics (desirable traits) from each of the parents. The offspring produced by sexual reproduction are not genetically identical to their parents or to each other; some offspring may be able to survive environmental changes better than either of their parents. However, one disadvantage of sexual reproduction is that some offspring with a different combination of traits may be less likely to survive than their parents. Nature is fine with this. The individual with desirable traits will have superior reproductive potential and help his/her species by passing a lot of good traits to the next generation. The individual with undesirable traits on the other hand will reproduce poorly or not at all and also help his/her species by not passing bad traits to the next generation. This gives rise to genetic diversity in sexually reproducing populations, which enables them to better adapt during the course of evolution. In other words, without crossover, we could only donate the exact copy of genes we inherited directly from our father or from our mother without shuffling them. Crossover is a mechanism to make sure our offspring get combinations of genes we have inherited by allowing swapping to get desirable or undesirable traits together.
The concept of crossover reminded me of a story between a very beautiful actress Sarah Bernhardt and George Bernard Shaw, Nobel Prize winner in Literature. When she asked, "Mr. Shaw, you and I should make love, for with my looks and your brains we would have wonderful children,” George Bernard Shaw responded, "Aha! But what if the child were born with my looks and your brain?" The crossover mechanism allows genes that are located in the chromosomes of different members to be combined under one chromosome, so that the offspring can inherit both of the desired genes or both of the undesired genes.
So, should we consider crossover as a mutation? The answer is No. In a mutation the final base sequence in the DNA differs from the base sequence in the original chromosomes. In a crossover, some parts of the chromosomes are swapped but unlike mutations, they are not deleted or changed, or new bases are added. However, those swapped parts of the DNA during crossover become a unique set of chromosome and inherited in the future generations. In other words, crossover allow mutant genes to be combined with each other. Without crossovers, if we had a desirable mutation on the paternal chromosome and a different desirable mutation on the maternal chromosome, we would never be able to get both desirable mutations in homozygous state. Therefore, we can conclude that without the mutation and the crossover, the evolution would essentially stop dead.
In birds, crossovers between the Z-chromosomes only occur in cocks during meiosis. Hens only have one Z-chromosome and crossover between the Z and W chromosome are impossible because of the heterochromatic character of these two chromosomes. However, hens can have crossover on autosomal chromosomes since they have 2 copies of every autosomal chromosomes. Although, we may not be able to see or notice the direct result of every crossover in the offspring’s phenotype, keep in mind that crossover happens all the time.
 In
pigeons, a good example of crossover would be qualmonds. However,
in order to understand the crossover from qualmond, we must first
understand the classical almond. A classical almond has a rich yellowish
ground color, just like the outside of the shell of an almond nut
from where the name was derived, and the body is sprinkled with dark
flecking, showing variegated effect with three distinct colors, black
(kite), ground (almond shell color), and white.
In
pigeons, a good example of crossover would be qualmonds. However,
in order to understand the crossover from qualmond, we must first
understand the classical almond. A classical almond has a rich yellowish
ground color, just like the outside of the shell of an almond nut
from where the name was derived, and the body is sprinkled with dark
flecking, showing variegated effect with three distinct colors, black
(kite), ground (almond shell color), and white.
There are 5 other allelic mutations to almonds which are faded, frosty, qualmond, sandy, and hickory. Qualmond mutation used to be known as blue almonds, and it was first discovered in a flock of barn pigeons in Portage County, Ohio in 1962 by Joe Quinn who named this mutation. At some point a wild-type mutated to almond. At some other time wild-type mutated to qualmond. Although both almond and qualmond mutations were totally independent events, qualmond is one of the alleles of almond and it is therefore located in the sex-chromosome. Qualmonds are generally a blue color with more or less flecking depending on the pattern as well as genes that enhance color like dirty. A T-pattern will typically be more flecked than a bar or barless. The main difference between the qualmond and almond is their base colors, where qualmond’s base color is blue, and qualmond breeding does not cause eye problems in the offspring unlike almonds. As an aside we can make qualmonds look just about like an almond in color. We simply need to add the kite, hetero recessive red and T pattern and darkeners. Generally qualmonds are less flecked than an almond, but not always. Like almonds, they fleck more with age.
When quamond is mated to brown birds, we are assuming there was a crossover between the b-locus genes for wild-type at brown and brown. As a result, the offspring were able to show brown based qualmond instead of the blue base. The very same thing must have happened when qualmond was mated to an ash-red. Because ash-red mutation is also located in the sex chromosome, a crossover made an otherwise blue based almond (qualmond) into an ash-red based. Therefore, due to crossover occurrence of qualmond with other sex-linked mutations, we can produce qualmond in all three combinations. However, since qualmond is a sex-linked gene, this particular crossover can only happen to cock birds since they have two alternatives of sex chromosomes (ZZ). In addition to the qualmond crossovers, ash red linked to almond and brown linked to almond have also been made. Crossover could occur in autosomal chromosomes as well. Ausomal mutations like Pattern (T-check, Check, Wild-type Bar, Barless), spread and recessive opal are located on one chromosome and presumably not very far from each other. Therefore we can have crossovers between these three genes. But they still all act on their own as individuals. The qualmond picture above is submitted by Vahe' D'Ala. www.falconlofts.com
Let me give you another example of crossover because it is a very important subject to understand fully. Lets say we have a blue bar cock bird that is +//+ at b locus and +//+ at the r locus; we mate him with a brown reduced hen (b//. and r//.). From the sex-linkage topic, we know that b locus (brown) and the r locus (reduced) are located on the sex chromosome. As expected, this mating produced a wild-type cock bird. Since he is a cock bird, we know he has two ZZ sex chromosomes, one he inherited from his blue bar father and one from his reduced brown mother. On the chromosome he inherited from his father he has wild-type at the b locus (B+) and wild-type (non-reduced) in the reduce locus (R+). On the chromosome that he inherited from his mother, he has brown mutation at the b-locus (b) and reduced mutation at the reduce locus (r). Therefore this cock bird presents himself as a blue bar in its phenotype, but we know he carries reduced and brown genes in his genotype. We also know that in the presence of the dominant allele, the recessive allele does not express itself. The only time that the recessive trait will be seen is when both of the alleles are recessive, b//b or r//r.
Now let's look at daughters this cock bird can produce. Because the genes under discussion are sex-linked, we are able to predict daughters' phenotypes. In the absence of crossover, this cock bird can produce daughters with only two different phenotypes. They could either be blue bar non-reduced (if he passes the chromosome he inherited from his father - B+, R+), or they could be brown and reduced (if he passes the chromosome he inherited from his mother - b, r). If however, this cock bird produce blue reduced (B+, r) daughters or brown non-reduced (b, R+) daughters, then we know that crossover has occurred.
Mendel's Laws of Inheritance
 Although
inheritance of biological traits has been recognized for thousands
of years, the first significant insights into the mechanisms involved
occurred in 1866, when Gregor Johann Mendel (right
picture) published the results of a series of experiments that would
lay the foundation
for the formal discipline of genetics. He was an Augustinian priest
and scientist, who gained posthumous fame as the figurehead of the
new science of genetics for his study of the inheritance of certain
traits in pea plants. Mendel's research was with plants, but the
basic underlying principles of heredity that he discovered also apply
to animals because the mechanisms of heredity are essentially the
same for all complex life forms. Mendel’s experiments proved
that some genes were inherited as units. From his study Mendel saw
that peas could be both short or tall plants, and they could be either
smooth or wrinkled when dried. By doing thousands of breeding tests
with pea plants, Mendel showed that these unit characters were transmitted
from parents to offspring unchanged. He also realized that some of
these characters were dominant or recessive to other units. In addition,
he saw that recessive character was not lost but carried in the gene
pool and recovered fully in future generations. Mendel had no idea
about genes, chromosomes, DNA, and the role and mechanism of meiosis
at that time and simply called these factors units of inheritance.
Mendel’s work produced two main principles of inheritance.
Although
inheritance of biological traits has been recognized for thousands
of years, the first significant insights into the mechanisms involved
occurred in 1866, when Gregor Johann Mendel (right
picture) published the results of a series of experiments that would
lay the foundation
for the formal discipline of genetics. He was an Augustinian priest
and scientist, who gained posthumous fame as the figurehead of the
new science of genetics for his study of the inheritance of certain
traits in pea plants. Mendel's research was with plants, but the
basic underlying principles of heredity that he discovered also apply
to animals because the mechanisms of heredity are essentially the
same for all complex life forms. Mendel’s experiments proved
that some genes were inherited as units. From his study Mendel saw
that peas could be both short or tall plants, and they could be either
smooth or wrinkled when dried. By doing thousands of breeding tests
with pea plants, Mendel showed that these unit characters were transmitted
from parents to offspring unchanged. He also realized that some of
these characters were dominant or recessive to other units. In addition,
he saw that recessive character was not lost but carried in the gene
pool and recovered fully in future generations. Mendel had no idea
about genes, chromosomes, DNA, and the role and mechanism of meiosis
at that time and simply called these factors units of inheritance.
Mendel’s work produced two main principles of inheritance.
One of these principles he found is now called Mendel's law of segregation. This law states that during the production of gametes (sperm and egg) the two copies of each hereditary factor segregate so that offspring acquire one factor from each parent, thereby ensuring that offspring have a "combination" of the parents' genes. For example, a pigeon has two allelic forms of a gene for pattern, one for checker and one for bar (wild-type) will transmit only one of these two alleles through a gamete to its offspring.
Mendel's second law of inheritance, The Law of Independent Assortment, also known as "Inheritance Law", states that alleles of different genes assort "independently" of one another during gamete formation. In other words, the chromosomes in gametes will be randomly selected from paternal or maternal chromosomes and will be passed to offspring. For example, on one homologous pair of chromosomes are the alleles for pattern (checker and bar) and on another pair of homologous are the alleles for grizzle and non-grizzle (wild-type). The segregation of alleles for pattern occurs independently of the segregation of the alleles for grizzle because each pair of homologoues behaves as an independent unit during meiosis. Furthermore, because the orientation of bivalents on the first meiotic metaphase plate is essentially random, four combinations of factors could be found in the meiotic products: (1) checker - grizzle, (2) checker - non-grizzle (wild-type), (3) bar (wild-type) - grizzle, (4) bar (wild-type) - non-grizzle (wild-type). This will insure that instead of nearly identical copies of an organism, a broad range of offspring develops, allowing more different abilities and evolutionary strategies for survival. This is very important for life to continue and create variety of individuals each time a new offspring is born.
We all have the same genes because we represent the human species, but we all have the different versions of the genes. We all look different because of these two laws of Mendel, and we all have different traits to carry and pass on to our offspring. The segregation is essential to why the traits do not "blend". The varying dominance of different genes causes some traits to appear unevenly instead of averaging out (whereby dominant doesn't mean more likely to reproduce - recessive genes can become the most common, too). In addition, segregation explains how recessive alleles that have no immediate effect on phenotype, can continue to be inherited for hundreds of generations. The Independent Assortment is essential for continued existence of "variation" but not a trending towards uniformity. These two laws of inheritance is also very important for pigeon breeders to understand the possible results when they breed their birds. This is how we could get brown pigmented birds from two ash-red parents, or a crested offspring from two non-crested parents.
Because of the "Inheritance
Law" two human brothers or sisters may look a bit alike but
will generally also be quite different. The only exception is identical
twins. In that case one single egg divides into two different cell
masses very, very early in the embryo and both cell masses wind up
forming a baby. Because the two embryos are the result of a single
egg/sperm combination, they
have the same genetic origins. Those two babies have identical genes as they
also have the same DNA.
The causes of identical twinning are generally unknown and unidentified.
No one really knows why an egg splits; technically it's a malfunction
of the normal development process. Birth rates for identical twins
are consistent across populations; it is the same regardless of race,
geography or maternal age. By the way, if you ever decided to ask
a parent of boy-girl twins if they are
identical twins, think again if you don’t want to look dumb.
Because all identical twins are always the same sex. As
far as we know there has never been a case of identical twins in pigeons
that actually hatched and survived. They simply die in the shell
because there seems to be no way for them to turn enough to crack the shell
and escape
at hatch time. There was
a study done by Hollander and Levi in 1940 “twins
and late embryonic monstrosities in pigeons”. Click
here to view this article.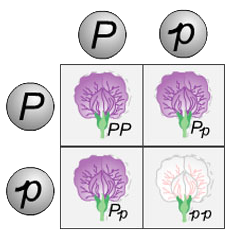
Punnet Square
The Punnett square is a diagram that is used to predict an outcome of a particular cross or breeding experiment. It is named after Reginald C. Punnett, who devised the approach, and is used by biologists to determine the probability of an offspring having a particular genotype. He discovered some basic principles of genetics, including sex linkage and sex determination. He worked with the feather color traits of chickens in order to quickly separate male and female chickens. The Punnett square is a summary of every possible combination of one maternal allele with one paternal allele for each gene being studied in the cross. The Punnett square validates Gregor Mendel's laws of genetic inheritance. We as a pigeon breeders need to be familiar with Punnett square, so we can calculate the possibilities of the expected phenotypes. Setting up and using a Punnett square is quite simple. You begin by drawing a grid of perpendicular lines, two rows and two columns if you are evaluating one trait. Next, you put the genotype of one parent across the top and that of the other parent down the left side. For example, if parent pea plant genotypes were Pp and Pp respectively. Note that only one letter goes in each box for the parents. It does not matter which parent is on the side or the top of the Punnett square. Lastly, all you have to do is fill in the boxes by copying the row and column-head letters across or down into the empty squares.
The Punnett square diagram demonstrated above for the flowers shows the mathematical probability of the possible genetic outcome when two heterozygous purple flowers (Pp) carrying white are crossed. In this case, white is recessive to purple and can only express itself in homozygous state (pp). Therefore, 25% or 1 out of 4 offspring from this mating should be a homozygous purple (PP). 50% or 2 out 4 offspring will be heterozygous purple carrying white, just like the phenotype and genotype of the parents (Pp). Finally, 25% or 1 out of 4 offspring should be a white flower (pp). The results from this cross clearly illustrate Mendel's principle of dominance and why the potential offspring traits appear with a predictable 3-to-1 dominant-to-recessive frequency.
You might be wondering why you are reading genetics about flowers in a pigeon genetics page. Because the careful breeding tests of pigeons clearly demonstrates that Mendel’s laws apply very well to pigeons. Some characteristics are however more complex than others and it is sometimes difficult to classify because of the exceptions to Mendel’s laws.
Exceptions to Simple Inheritance
Our knowledge of genetic inheritance grown immensely since Gregor Mendel and we now know certain inheritance rules have exceptions. Mendel happened to use pea plants, which happened to have a number of easily observable traits that were determined by just two alleles. The traits Mendel studied in his peas, one allele happened to be dominant for the trait & the other was a recessive form. In Mendel’s classic pea crosses, the offspring always looked like one of the two parental varieties because one allele in a pair showed complete dominance over the other. In such situations, the phenotypes of heterozygote and the dominant homozygote are indistinguishable. However, things aren't always so clear-cut & "simple" in the world of genetics. Genetic Science, with the help of technology now have a lot more complex information, adding discoveries to Mendel’s ingenious work.
In 1910, Thomas Hunt Morgan at
Columbia University discovered crossover (recombinant genes). This
suggested that there was linkage
of genes but, if linkage exists, why didn't Mendel detect it while
carrying out his crosses in pea plants? Well, Mendel only studied
seven traits which were all controlled by a single gene in pea
plants. Pea plants also have only seven chromosomes. Still, Mendel
didn't
choose pairs of genes that were always on different chromosomes;
some of the pairs of genes that Mendel studied were actually on
the same chromosomes. However, because the genes on these chromosomes
are so distantly located from each other, they behave as though
they
independently assort. Thus, Mendel was unable to detect linkage.
The trait that controlled the smooth vs. wrinkled (V) seeds and
the trait that controlled the height of the plant (Le) are located
on
the same chromosome. Hence, if Mendel had completed the cross,
possibly with just one more cross, he would have discovered linkage
himself.
There were other exceptions to Mendel's inheritance laws. One of these exceptions is called Incomplete dominance which is a condition when neither allele is dominant and recessive over the other, like the ones Mendel studied in his pea plants. Some alleles are equally strong and neither are masked by the other and when crossed both alleles are expressed in the phenotype. The hybrid is a blend of both alleles resulting in an intermediate expression of a trait in heterozygous individuals. In some plants, if a red flowered plant is crossed with a white flowered one, the progeny will all be pink. It's like mixing paints, red + white will make pink. The pink flowers result because the single "red" allele is unable to code for the production of enough red pigment to make the petals dark red. In addition, when pink flowered plant is crossed with another pink flowered plant, the progeny are 1 red, 2 pink, and 1 white. Indigo mutation in pigeons would be a good example of incomplete dominance.
 The
other exception to Mendels inheritance law is called co-dominance which
is a situation in which both alleles are equally strong and both
alleles are visible in the hybrid's phenotype. Co-dominance only
applies to different alleles of the same gene as all forms of dominance
do. The
genetic
gist to co-dominance is pretty much the same as incomplete dominance.
A
hybrid organism shows a third phenotype --- not the usual "dominant" one & not
the "recessive" one, but a third, different phenotype. In co-dominance,
the "recessive" & "dominant" traits appear together in the phenotype
of the hybrid organisms. The
flower on the left is a good example of a co-dominance hybrid where
we see both
alleles (red and white) in the hybrid's phenotype. Grizzle mutation in
pigeons would be a good example of co-dominance.
The
other exception to Mendels inheritance law is called co-dominance which
is a situation in which both alleles are equally strong and both
alleles are visible in the hybrid's phenotype. Co-dominance only
applies to different alleles of the same gene as all forms of dominance
do. The
genetic
gist to co-dominance is pretty much the same as incomplete dominance.
A
hybrid organism shows a third phenotype --- not the usual "dominant" one & not
the "recessive" one, but a third, different phenotype. In co-dominance,
the "recessive" & "dominant" traits appear together in the phenotype
of the hybrid organisms. The
flower on the left is a good example of a co-dominance hybrid where
we see both
alleles (red and white) in the hybrid's phenotype. Grizzle mutation in
pigeons would be a good example of co-dominance.
Most scientist and
scientific writings will consider co-dominance
and incomplete dominance as the
dominances of a lesser degree. In fact, they will use co-dominance as
the preferred term for both conditions because it is the simplest, shortest,
and the most adequate term.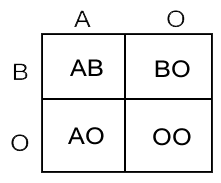
The other good example of co-dominance would be the blood type in humans. The ABO blood group system is widely credited to have been discovered by the Austrian scientist Karl Landsteiner, who found three different blood types in 1900; he was awarded the Nobel Prize in Physiology or Medicine in 1930 for his work. The ABO blood group is a major system for classifying blood types in humans. Blood type AB is inherited in a co-dominant pattern. If a father with blood type AO (type A heterozygous for type O) and a mother with blood type BO (type B heterozygous for type O) have four children, each child could be with a different blood type: AO, AB, BO, and OO. Therefore, the ABO blood type system is also an exception of a trait that is controlled by more than just a single pair of alleles which is not what Mendel’s work with pea plants have suggested. In other words, it is due to a multiple-allele series instead of two. In this case, there are three alleles (A, B, and O), but each individual only inherits two of them, one from each parent. We currently don’t have any data about the blood type system of pigeons.
Co-dominance in pigeons
In pigeons, a mutant gene is said to be co-dominant when a bird that has one copy (heterozygous) of that mutant and one copy of the wild-type gene at that locus and looks different than wild-type. In addition, if the bird has two copies (homozygous) of that mutant gene it will show a second distinct phenotype. In pigeons, we can confidently say that most genes are actually co-dominant but we can only see some of them as clear examples when we look at their phenotype. It is very important to understand that, there is no bright line distinction between co-dominant and dominant mutations. According to Dr. Cryberg, there are not many true dominants in pigeons. Spread probably comes about as close to dominant as anything. Yet, some heterozygous spread is not as black as a homozygous spread. Check and T-pattern are other examples much like spread. A bird heterozygous for check and bar will not have as large checks compared to a bird that is homozygous for check. But in both cases of spread and check they are close enough to be fairly called dominants.
The line between a recessive and co-dominant or between co-dominant and dominant is not always clear in genetic science. There are lots of genes that are very close to recessive. But if you look very close, you can often see hints on wild-type that they are there even in heterozygous state. The same is true of co-dominant vs. dominant genes. Let’s take spread as an example which is usually listed as simply a dominant mutation. Yet, a heterozygous spread blue bar is not near as black as a homozygous spread blue bar. This concludes that not everything in pigeon genetics is 100% dominant or recessive to wild-type. In fact, we could say that in pigeons anything that is 90% recessive is generally called simply recessive and anything that is 90% dominant is simply called dominant. Thus, everything in between should be considered as co-dominant if that will help you understand the argument better between dominant, recessive, and co-dominant genes.
 In
pigeons, grizzle, almond, indigo and dominant opal are fair examples
of co-dominant genes where
there are two distinct phenotypes clearly visible based on their heterozygous
or homozygous state compared to wild-type.
In
pigeons, grizzle, almond, indigo and dominant opal are fair examples
of co-dominant genes where
there are two distinct phenotypes clearly visible based on their heterozygous
or homozygous state compared to wild-type.
For example, a grizzle mutation in heterozygous state on an otherwise wild-type pigeon will express black and white feathers (salt and pepper phenotype with the blue/black color and white mixed) all over the pigeon's body. The amount of black and white feathers is determined by the pattern of the grizzle birds. If the bird is T-checker for pattern for example, white feathers on the wings would be less than a bird with a bar pattern. The grizzle mutation in heterozygous state also has a tiger grizzle phenotype, where white feathers randomly mixed with colored feathers. So, heterozygous grizzles show two different phenotypes than wild-type as normal grizzle and tiger grizzle. Same grizzle mutation in homozygous state however, will make the same bird look like a solid white bird except only the very ends or tips of its wing and tail feathers would be pigmented. Most fanciers would call this phenotype a stork marking (The Iranian High Flyer picture on the left demonstrates this phenotype). Therefore, we can say that grizzle mutation in pigeons will show lesser co-dominant effect in heterozygous state than homozygous state. Because grizzle mutation shows three distinct phenotypes depending on its absence (wild-type), its presence as a heterozygote (salt and pepper phenotype) or its presence as homozygote (stork marking), by definition grizzle is a co-dominant mutation.
In the case of almond, we can only talk about cock birds as almond is a sex-linked mutation and therefore located in the sex chromosome. Because hens only have one functional sex chromosome, they cannot have two copies of a sex-linked gene. Thus, a heterozygous almond cock will have the phenotype of a normal almond. If the cock bird is homozygous for the almond mutant, then this bird, just like in homozygous grizzle example would present itself as near white, a second distinct phenotype. This makes almond another example of co-dominant mutation in pigeons.
A heterozygous dominant opal is somewhat bronzed usually and somewhat lighter in color than wild-type. Breeding data suggest that homozygous dominant opal is usually lethal. The few suspected homozygous dominant opals that lived showed near white phenotype, and they had abnormal feathers and were short lived. Again, because dominant opal mutation clearly shows second distinct phenotype depending on its heterozygous vs. homozygous (lethal) state, it is also a co-dominant mutation.
A heterozygous indigo simply bronzes the bird a bit and also nearly removes the tail bar. A homozygous indigo on the other hand looks like an ash red mimic where the tail has no sub-terminal tail bar. Therefore, indigo mutation presenting two distinct phenotypes depending on heterozygous vs. homozygous state is also well known examples of a co-dominance mutation found in pigeons.
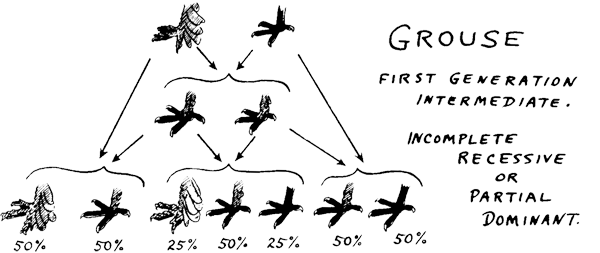 The
picture on the right is taken from N.P.A Information Booklet #1, Project
on Genetics. Grouse in pigeons is one of the examples
used to explain the co-dominance condition. If we cross a clean-legged
pigeon to a pigeon with feathered feet, first generation will show
co-dominance where the offspring will have feathers on its legs.
However, length of the feet feathers will not be as long as the parent
with feathered legs. Instead, it will be intermediate in length.
Crossing the first generation with the parents will result in an
incomplete recessive or partial dominance in the second generation.
The
picture on the right is taken from N.P.A Information Booklet #1, Project
on Genetics. Grouse in pigeons is one of the examples
used to explain the co-dominance condition. If we cross a clean-legged
pigeon to a pigeon with feathered feet, first generation will show
co-dominance where the offspring will have feathers on its legs.
However, length of the feet feathers will not be as long as the parent
with feathered legs. Instead, it will be intermediate in length.
Crossing the first generation with the parents will result in an
incomplete recessive or partial dominance in the second generation.
Slipper is another mutation creates muffs in legs, and it is also co-dominant as the grouse. The hetero versions of both grouse and slipper produce birds that have either feathers or scale pattern disruptions on the shanks and toes that can be seen easy enough. Thus, in both mutations the heterozygote differs from both wild-type and from the homozygote to produce second different phenotype and that means they are co-dominants. Click here to read an excellent article written by Robert J. Mangile about Synthetic Muffs in Pigeons.
Epistatis & Supplementary Mutations
There
is this other condition known as epistasis,
where one gene suppresses the effects of another non-allelic gene. In
other words, when two dominant
genes are present but only one of them is expressed on the phenotype,
then the gene that is expressed is said to be epistatic. Hypostasis,
on the other hand is the converse, and is applied to the gene pair hidden
by the epistatic gene pair. If
we have a pigeon that is spread but is also a checker for pattern, this
bird will not be able to express the check pattern on the phenotype
because of the spread gene. A Spread on a blue bird makes an otherwise
blue pigeon a solid black pigeon and shows no evidence of pattern. Same
is true for all other patterns where spread gene conceals (hides) all the
possible patterns, T-Check, Check, Bar, and Barless. Therefore, spread
is said to be epistatic to all known patterns. Recessive
Red is another good example of epistatis condition because in homozygous
state a recessive red will conceal most other colors (except recessive
white and albino) and patterns. That is, it "covers" color
so that, whether the bird is ash-red or blue-black, it appears as recessive red.
It likewise covers the pattern, so you can't tell whether the bird is check or
bar, beneath the recessive red. Epistatis
differs from dominance in that it is not on the same chromosome as the
mutation it hides. For example
recessive white hides all other color mutants and
it can correctly said to be epistatic to all other known color mutants,
except
homozygous albino.
When we look at a recessive white bird, we would not have any clue what
other
color genes and patterns might be present in its genotype. So
to recap quickly, the albino
is a recessive autosomal mutation and is known to be epistatic to “all” known
color and pattern mutations. Recessive white is epistatic to all
know colors and patterns, except albino. Recessive red is epistatic
to all patterns and most color mutants, except recessive white, and
albino. And finally, spread is epistatic to all known pattern series
T-check, Check, Bar and Barless.
The second way epistasis
is used is a bit tricky. In this case wild-type can be epistatic
to some mutant; however, this mutant can show if some other mutant
is present. An example would be white sides on recessive red. A recessive
red white sides bird is a self red bird with white wing shields.
Yet, if you simply replace the recessive red mutant with wild-type
at that locus you will get a self blue bird with no white showing
even when the mutant for white sides is homozygous. So wild-type
is said to be epistatic to the white sides mutant. White sides is
neither
dominant, nor recessive, nor codominant. It simply does not show
on an otherwise wild-type pigeon. There are other known examples
where wild-type is epistatic to some mutant. In fact it is not all
that uncommon. But very few pigeon people recognize the situation Dr.
Richard Cryberg (private communication). 
Another well recognize instance is the white wing mutation as found in Archangel breed. White wing is only expressed in the presence of the ash-red mutation and it will not show on wild-type, brown and recessive red. Notice that we are now talking about “white wing” mutation that we normally see in ash-red phenotype. The “white sides” that we normally see in recessive red is a different gene than “white wings”. Both genes produce similar phenotypes where a self red bird presents itself with the white wing shields. The only difference is that white sides only shows up in recessive red, and the white wing only shows up in ash-red. It is important to notice that these genes only present themselves in red plumage and not with brown or black. Although a lot more testing is needed, our current understanding suggest that white wing acts like a sex-linked trait. Just like the “white side”, wild-type is also epistatic to the “white wing”.
There are many known examples of wild-type being epistatic to mutations in both animals, birds, and plants. These types of mutations are not particularly uncommon in species that have been long bred for particular "fancy" phenotypes. Such mutations have a scientific name. They are called supplementary mutations. The word supplementary packs a lot of meaning in a single word. The first thing the word tells you is that wild-type is epistatic to the mutation under discussion. The mutant does not show on an otherwise wild-type individual even if the mutation is homozygous. The second thing the word supplementary tells you is that there exists a known mutation that will allow the supplementary mutant to show phenotypically. It may even show when the supplementary mutant is heterozygous. So both white sides on recessive red and white wing on ash-red are properly called supplementary mutants. There is no question that supplementary genes exist in pigeons beyond the two listed examples based on unpublished studies carried out by Dr. Cryberg (private communication).
So, we have two different supplementary mutants (white side and white wing) that both produce white wing shields. But the one that works on ash red does not work on recessive red. And the one that works on recessive red does not work on ash red. When describing such mutations the short way to explain the behavior would be to say that white sides is a supplementary mutant that is codominant to homozygous recessive red. Note - this is the only case in genetics where it is proper to rate dominance, codominance or recessiveness versus something other than either wild-type or another allele of the mutant. There is also another way to express this type of inheritance. We could say that wild-type is epistatic to the white sides mutant but that this mutant expresses as a codominant in the presence of homozygous recessive red.
Pleiotropy
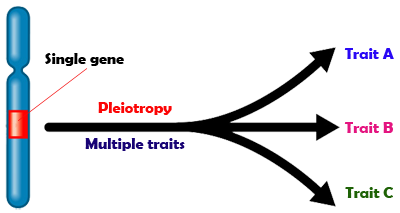 So
far, we have treated Mendelian inheritance as though each gene affects
only one phenotypic character. There is another exception
to Mendel’s inheritance law: pleiotropy, a condition in which
a single gene influences more than one phenotypic
trait in an organism. In other words, pleiotropism is the control
by a single gene of several distinct and seemingly unrelated phenotypic
effects. Consequently, a mutation in a pleiotropic gene may have
an effect on some or all traits simultaneously. Pleitropy has been
identified in a wide range of species, including humans. The word
pleiotropy has a Greek origin; it comes from the words pleion, meaning "more," and
trepein, meaning "to turn" or "to convert." The
pleiotropy concept is not generally covered in pigeon genetics books,
but in my opinion it is an important concept to understand. When
studying genetics, because most mutants cause some degree of pleiotropy,
it is something to be expected, not something that is surprising.
Pleiotropy should not be confused with polygenic traits, in which
multiple genes converge to result in a single phenotype e.g., Toy
Stencil and Frill Stencil in pigeons.
So
far, we have treated Mendelian inheritance as though each gene affects
only one phenotypic character. There is another exception
to Mendel’s inheritance law: pleiotropy, a condition in which
a single gene influences more than one phenotypic
trait in an organism. In other words, pleiotropism is the control
by a single gene of several distinct and seemingly unrelated phenotypic
effects. Consequently, a mutation in a pleiotropic gene may have
an effect on some or all traits simultaneously. Pleitropy has been
identified in a wide range of species, including humans. The word
pleiotropy has a Greek origin; it comes from the words pleion, meaning "more," and
trepein, meaning "to turn" or "to convert." The
pleiotropy concept is not generally covered in pigeon genetics books,
but in my opinion it is an important concept to understand. When
studying genetics, because most mutants cause some degree of pleiotropy,
it is something to be expected, not something that is surprising.
Pleiotropy should not be confused with polygenic traits, in which
multiple genes converge to result in a single phenotype e.g., Toy
Stencil and Frill Stencil in pigeons.
Take albinism as an example, which is a very common mutation in the animal kingdom that eliminates pigment production. The albino mutation interferes with the production of melanin, none is produced anywhere in the body. Albinism is a pleiotropic trait where the gene for this trait not only results in the typical albino deficiency of skin, hair, and eye pigmentation, but also causes defects in vision. Therefore, an albino pigeon will have pink eyes, white feathers, an ivory colored beak, and nails; thus, one gene is influencing more than one phenotypic trait.
Another example of pleiotropy in pigeons is false pearl eye produced on brown pigmented pigeons. According to Joe Quinn, brown pigeons seldom have orange eyes because the brown mutation affects certain aspect of iris pigmentation by producing a creamy white “pearl-eye” like coloration, which is known as the “false pearl” eyes. According to Dr. Cryberg, the usual function of the wild-type at brown gene is to make an enzyme that allows oxidation of a particular compound that makes black pigment when oxidized. However, in the case of brown mutation, the gene that makes the black pigment (wild-type) is mutated, so oxidation does not happen; therefore, a brown pigment is produced instead of black. “In pigeon eyes there are two non melanin pigments. One is called a guanidine and the other a pteridine. The guanidine makes a whitish opaque pigment while the pteridine makes an orange pigment. Apparently the chemical produced by wild type at brown is not only important in allowing production of black pigment but also important in making orange pteridine. The reason is brown pigeons do not seem to make any pteridine and as a result have false pearl eyes” says Dr. Cryberg.
Dominant Opal (Od) is another good example of pleiotropy in pigeons. We know heterozygous Od birds show diluted color and they are more or less bronzed. Od also whitens the pattern and sub-terminal tail band in hetero state. In the homozygous state however, Od birds show near white phenotype, and, by definition, we call this gene co-dominant since it shows a second distinct phenotype (different plumage color) in homozygous state. We also know that in homozygous state Od is lethal. But, from a lethal standpoint, a hetero Od bird is perfectly normal or so close to perfectly normal that no one has ever reported otherwise. Therefore, the lethal trait in Od bird is not co-dominant; it is recessive since it is only lethal in homozygous state. Thus, the homozygous Od bird dies before hatching most of the time. It seldom hatches, but when it does, it never lives to sexual maturity. Therefore, it is perfectly fair to say that Od is a co-dominant color mutant and a recessive lethal mutant. Since the same gene (Od) causes color change as well as potentially lethal effects, Od is also a good example of a pleiotropy in pigeon genetics.
In addition to the examples given above, according to Dr. Cryberg, a lot of other mutants are also pleiotropic in pigeons:
Dilute – This mutation dilutes the plumage color; all of the newly hatched dilute babies have short down length, and ease of plucking squabs. Many years ago, Levi showed that in his commercial squabbers, dilutes did not produce as many young per year as intense birds. This was a disappointment as the women he had that plucked feathers, much preferred plucking dilutes, as they could do them faster.
Recessive white – This mutation turns the plumage color to white, no matter what other color mutant genes are present in the genotype. It also changes the eye color to black, (bull eye), again concealing the otherwise orange or pearl eye colors.
Almond – This mutation dilutes plumage color, but leaves normal intensity flecks; furthermore, there are vision issues in a homozygous bird. Since almond is a sex-linked mutation, only cock birds can be homozygous for this trait, and only cock birds have eye problems in the homozygous state.
Recessive red – In this mutation, the plumage color changes from the normal blue/black to red, and it greatly enhances the molt to white tendency when feathers are plucked repeatedly. In addition, recessive red also turns wild-type to red, except in some strains of Homers where the bird is mainly red, but also shows random feathers that are as black as can be. This is very unlikely to be either mosaic or chimeric as it happens to every bird from many pairs. The color is not patches. It is random feathers here and there.
There are a lot more exceptions to Mendelian inheritance which refers to the expression of inheritance of monogenic traits, traits that are controlled by one gene. I think the information covered above is enough material for basic genetics and genetic terms for now. It was imperative for me to mention all the technical words and explain basic genetics above, so that you could understand the rest of the material in next pages. You probably noticed self-learning by reading is not as easy as you think. Most people read and comprehend their reading much more slowly than listening, watching, or experimenting. You can quickly read anything but understanding what you read takes time. Just because you read it doesn't mean you understand it either. Reading and scrolling down the screen is no way to learn (anything). You must interact with the reading. I highly recommend that you go back and re-read the material above if you have to, read at your own pace and remember that you are here to learn genetics not to simply become a geneticist.
Now, let’s look at the history of pigeons, their interaction with human and their mutations through natural selection and evolution.